

NRF1 association with AUTS2-Polycomb mediates specific gene activation in the brain
- PMID: 34637754
- PMCID: PMC8604784
- DOI: 10.1016/j.molcel.2021.09.020
NRF1 association with AUTS2-Polycomb mediates specific gene activation in the brain
Erratum in
-
NRF1 association with AUTS2-Polycomb mediates specific gene activation in the brain.Mol Cell. 2021 Nov 18;81(22):4757. doi: 10.1016/j.molcel.2021.10.023. Mol Cell. 2021. PMID: 34798045 No abstract available.
Abstract
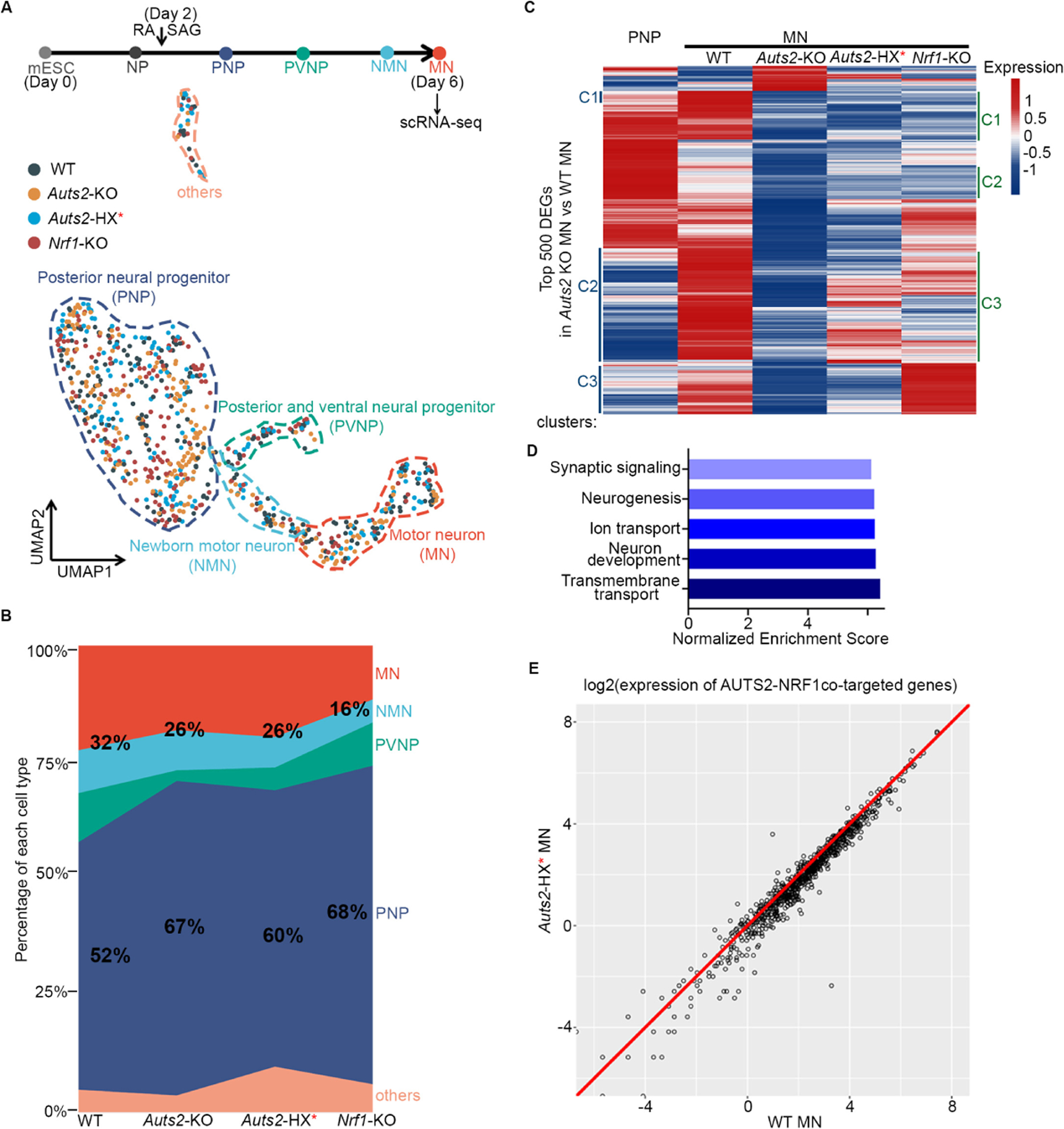
The heterogeneous family of complexes comprising Polycomb repressive complex 1 (PRC1) is instrumental for establishing facultative heterochromatin that is repressive to transcription. However, two PRC1 species, ncPRC1.3 and ncPRC1.5, are known to comprise novel components, AUTS2, P300, and CK2, that convert this repressive function to that of transcription activation. Here, we report that individuals harboring mutations in the HX repeat domain of AUTS2 exhibit defects in AUTS2 and P300 interaction as well as a developmental disorder reflective of Rubinstein-Taybi syndrome, which is mainly associated with a heterozygous pathogenic variant in CREBBP/EP300. Moreover, the absence of AUTS2 or mutation in its HX repeat domain gives rise to misregulation of a subset of developmental genes and curtails motor neuron differentiation of mouse embryonic stem cells. The transcription factor nuclear respiratory factor 1 (NRF1) has a novel and integral role in this neurodevelopmental process, being required for ncPRC1.3 recruitment to chromatin.
Keywords: AUTS2; NRF1; P300; RSTS; active transcription; brain development; ncPRC1.3; polycomb.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests D.R. is a cofounder of Constellation and Fulcrum Pharmaceuticals.
Figures






References
-
- Ajmone PF, Avignone S, Gervasini C, Giacobbe A, Monti F, Costantino A, Esposito S, Marchisio P, Triulzi F, and Milani D (2018). Rubinstein-Taybi syndrome: New neuroradiological and neuropsychiatric insights from a multidisciplinary approach. Am J Med Genet B Neuropsychiatr Genet 177, 406–415. - PubMed
-
- Aloia L, Stefano BD, and Croce LD (2013). Polycomb complexes in stem cells and embryonic development. Development 140, 2525–2534. - PubMed
-
- Bannister AJ, and Kouzarides T (1996). The CBP co-activator is a histone acetyltransferase. Nature 384, 641–643. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
