

Single-cell sequencing reveals antitumor characteristics of intratumoral immune cells in old mice
- PMID: 34642245
- PMCID: PMC8513495
- DOI: 10.1136/jitc-2021-002809
Single-cell sequencing reveals antitumor characteristics of intratumoral immune cells in old mice
Abstract
Background: Aging has long been thought to be a major risk factor for various types of cancers. However, accumulating evidence indicates increased resistance of old animals to tumor growth. An in-depth understanding of how old individuals defend against tumor invasion requires further investigations.
Methods: We revealed age-associated alterations in tumor-infiltrating immune cells between young and old mice using single-cell RNA and coupled T cell receptor (TCR) sequencing analysis. Multiple bioinformatics methods were adopted to analyze the characteristics of the transcriptome between two groups. To explore the impacts of young and old CD8+ T cells on tumor growth, mice were treated with anti-CD8 antibody every 3 days starting 7 days after tumor inoculation. Flow cytometry was used to validate the differences indicated by sequencing analysis between young and old mice.
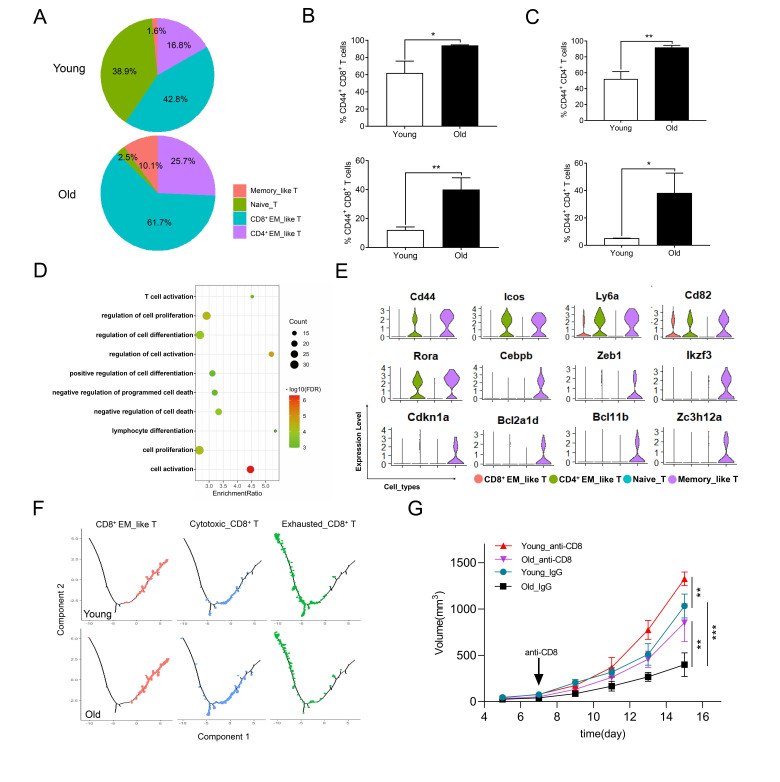
Results: We found a higher proportion of cytotoxic CD8+ T cells, naturally occurring Tregs, conventional dendritic cell (DC), and M1-like macrophages in tumors of old mice compared with a higher percentage of exhausted CD8+ T cells, induced Tregs, plasmacytoid DC, and M2-like macrophages in young mice. Importantly, TCR diversity analysis showed that top 10 TCR clones consisted primarily of exhausted CD8+ T cells in young mice whereas top clones were predominantly cytotoxic CD8+ T cells in old mice. Old mice had more CD8+ T cells with a 'progenitor' and less 'terminally' exhausted phenotypes than young mice. Consistently, trajectory inference demonstrated that CD8+ T cells preferentially differentiated into cytotoxic cells in old mice in contrast to exhausted cells in young mice. Importantly, elimination of CD8+ T cells in old mice during tumor growth significantly accelerated tumor development. Moreover, senescent features were demonstrated in exhausted but not cytotoxic CD8+ T cells regardless of young and old mice.
Conclusions: Our data revealed that a significantly higher proportion of effector immune cells in old mice defends against tumor progression, providing insights into understanding the altered kinetics of cancer development and the differential response to immunotherapeutic modulation in elderly patients.
Keywords: CD8-positive T-lymphocytes; cellular; immunity; immunotherapy; lymphocytes; tumor microenvironment; tumor-infiltrating.
© Author(s) (or their employer(s)) 2021. Re-use permitted under CC BY-NC. No commercial re-use. See rights and permissions. Published by BMJ.
Conflict of interest statement
Competing interests: None declared.
Figures







References
-
- Shanmugham LN, Petrarca C, Frydas S, et al. . Il-15 an immunoregulatory and anti-cancer cytokine. recent advances. J Exp Clin Cancer Res 2006;25:529–36. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Medical
Research Materials