

Cyclic CMP and cyclic UMP mediate bacterial immunity against phages
- PMID: 34644530
- PMCID: PMC9070634
- DOI: 10.1016/j.cell.2021.09.031
Cyclic CMP and cyclic UMP mediate bacterial immunity against phages
Abstract
The cyclic pyrimidines 3',5'-cyclic cytidine monophosphate (cCMP) and 3',5'-cyclic uridine monophosphate (cUMP) have been reported in multiple organisms and cell types. As opposed to the cyclic nucleotides 3',5'-cyclic adenosine monophosphate (cAMP) and 3',5'-cyclic guanosine monophosphate (cGMP), which are second messenger molecules with well-established regulatory roles across all domains of life, the biological role of cyclic pyrimidines has remained unclear. Here we report that cCMP and cUMP are second messengers functioning in bacterial immunity against viruses. We discovered a family of bacterial pyrimidine cyclase enzymes that specifically synthesize cCMP and cUMP following phage infection and demonstrate that these molecules activate immune effectors that execute an antiviral response. A crystal structure of a uridylate cyclase enzyme from this family explains the molecular mechanism of selectivity for pyrimidines as cyclization substrates. Defense systems encoding pyrimidine cyclases, denoted here Pycsar (pyrimidine cyclase system for antiphage resistance), are widespread in prokaryotes. Our results assign clear biological function to cCMP and cUMP as immunity signaling molecules in bacteria.
Keywords: Pycsar; anti-phage; bacteria; cCMP; cUMP; cyclase; defense; pb8; phage; pyrimidine.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests R.S. is a scientific cofounder and advisor of BiomX, Pantheon Bioscience, and Ecophage.
Figures


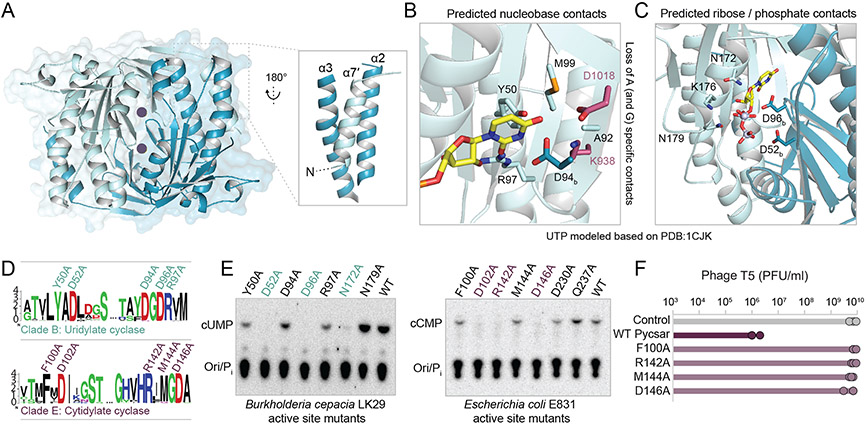

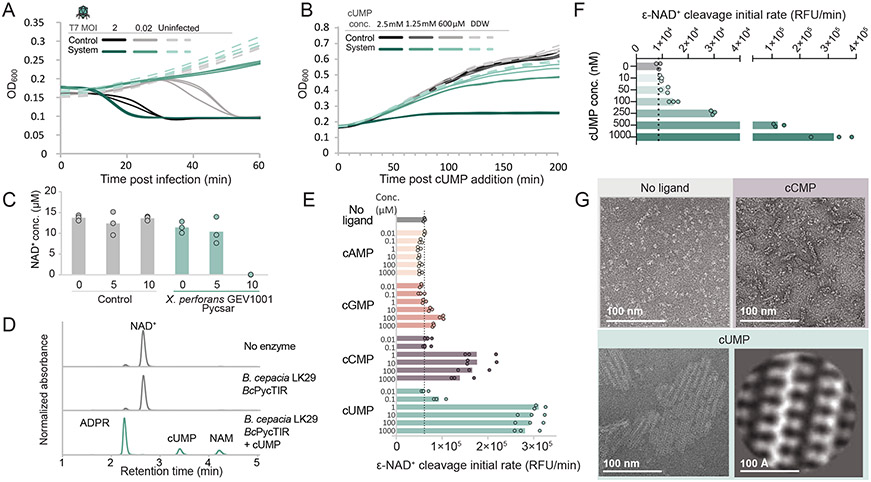

Comment in
-
Cyclic pyrimidines defend against phages.Nat Rev Microbiol. 2022 Jan;20(1):1. doi: 10.1038/s41579-021-00658-w. Nat Rev Microbiol. 2022. PMID: 34697499 No abstract available.
-
Cyclic pyrimidines jump on the anti-phage bandwagon.Cell. 2021 Nov 11;184(23):5691-5693. doi: 10.1016/j.cell.2021.10.012. Cell. 2021. PMID: 34767773
References
-
- Bähre H, Danker KY, Stasch JP, Kaever V, and Seifert R (2014). Nucleotidyl cyclase activity of soluble guanylyl cyclase in intact cells. Biochem. Biophys. Res. Commun 443, 1195–1199. - PubMed
-
- Beckert U, Grundmann M, Wolter S, Schwede F, Rehmann H, Kaever V, Kostenis E, and Seifert R (2014). cNMP-AMs mimic and dissect bacterial nucleotidyl cyclase toxin effects. Biochem. Biophys. Res. Commun 451, 497–502. - PubMed
-
- Bernheim A, and Sorek R (2020). The pan-immune system of bacteria: antiviral defence as a community resource. Nat. Rev. Microbiol 18, 113–119. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
