

The cholesterol metabolite 25-hydroxycholesterol restrains the transcriptional regulator SREBP2 and limits intestinal IgA plasma cell differentiation
- PMID: 34644558
- PMCID: PMC8570345
- DOI: 10.1016/j.immuni.2021.09.004
The cholesterol metabolite 25-hydroxycholesterol restrains the transcriptional regulator SREBP2 and limits intestinal IgA plasma cell differentiation
Abstract
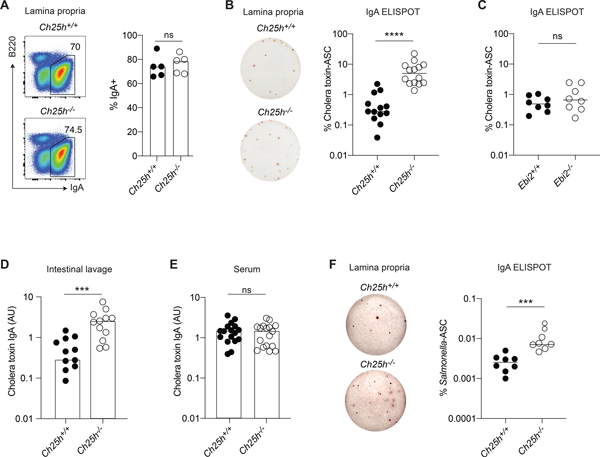
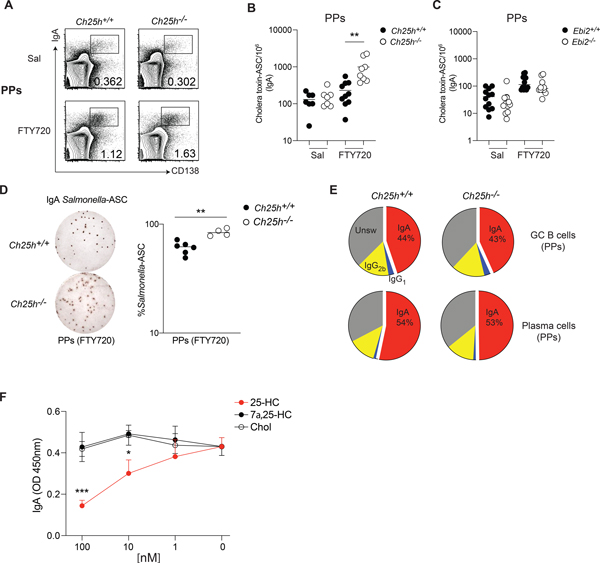
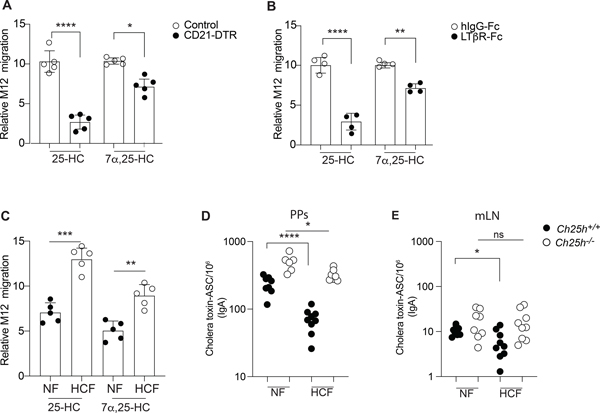
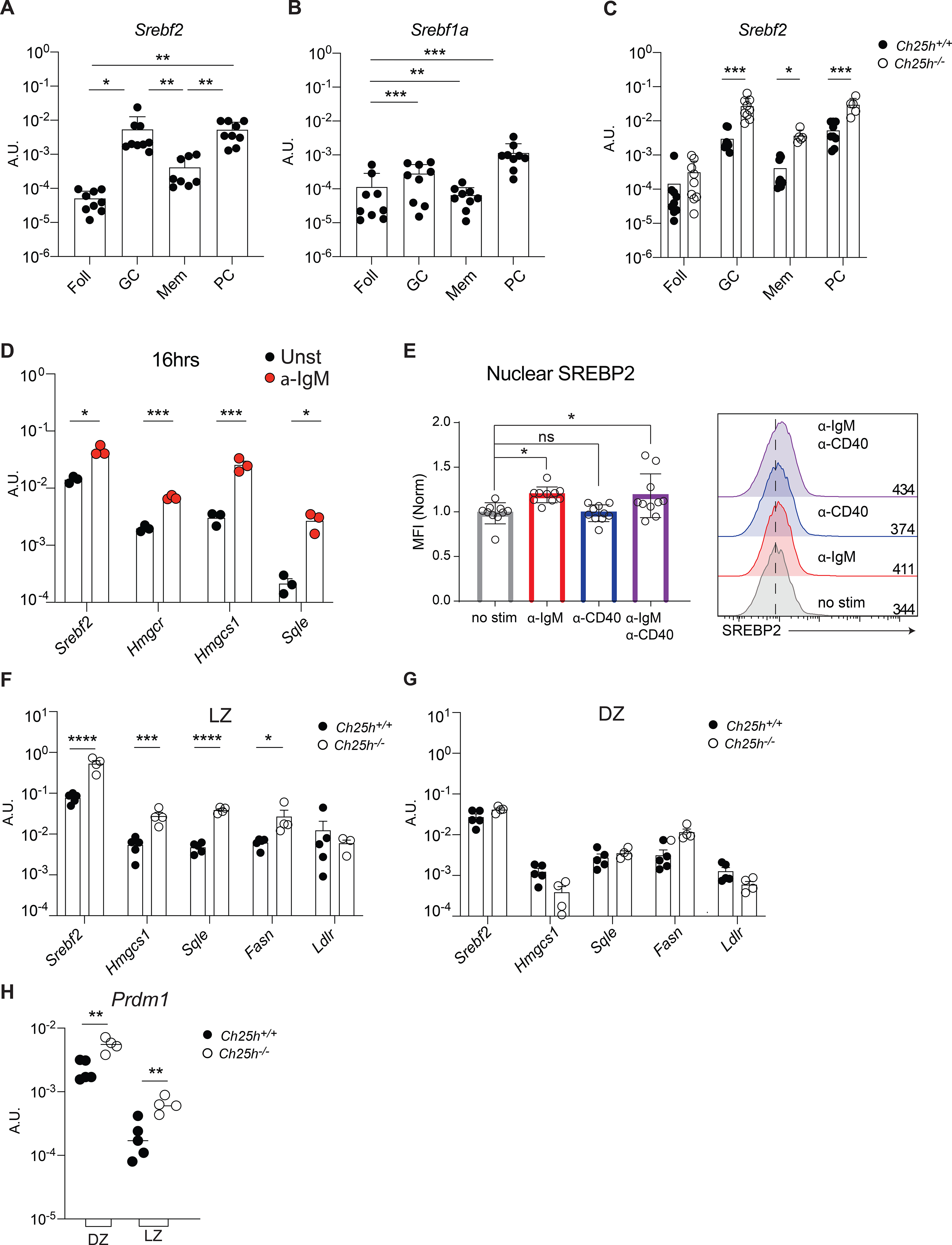
Diets high in cholesterol alter intestinal immunity. Here, we examined how the cholesterol metabolite 25-hydroxycholesterol (25-HC) impacts the intestinal B cell response. Mice lacking cholesterol 25-hydroxylase (CH25H), the enzyme generating 25-HC, had higher frequencies of immunoglobulin A (IgA)-secreting antigen-specific B cells upon immunization or infection. 25-HC did not affect class-switch recombination but rather restrained plasma cell (PC) differentiation. 25-HC was produced by follicular dendritic cells and increased in response to dietary cholesterol. Mechanistically, 25-HC restricted activation of the sterol-sensing transcription factor SREBP2, thereby regulating B cell cholesterol biosynthesis. Ectopic expression of SREBP2 in germinal center B cells induced rapid PC differentiation, whereas SREBP2 deficiency reduced PC output in vitro and in vivo. High-cholesterol diet impaired, whereas Ch25h deficiency enhanced, the IgA response against Salmonella and the resulting protection from systemic bacterial dissemination. Thus, a 25-HC-SREBP2 axis shapes the humoral response at the intestinal barrier, providing insight into the effect of high dietary cholesterol in intestinal immunity.
Keywords: 25-hydroxycholesterol; Peyer's patches; dietary cholesterol; follicular dendritic cells; immunoglobulin A; plasma cells; sterol response element binding proteins 2.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declarations of interest The authors declare no competing interests.
Figures
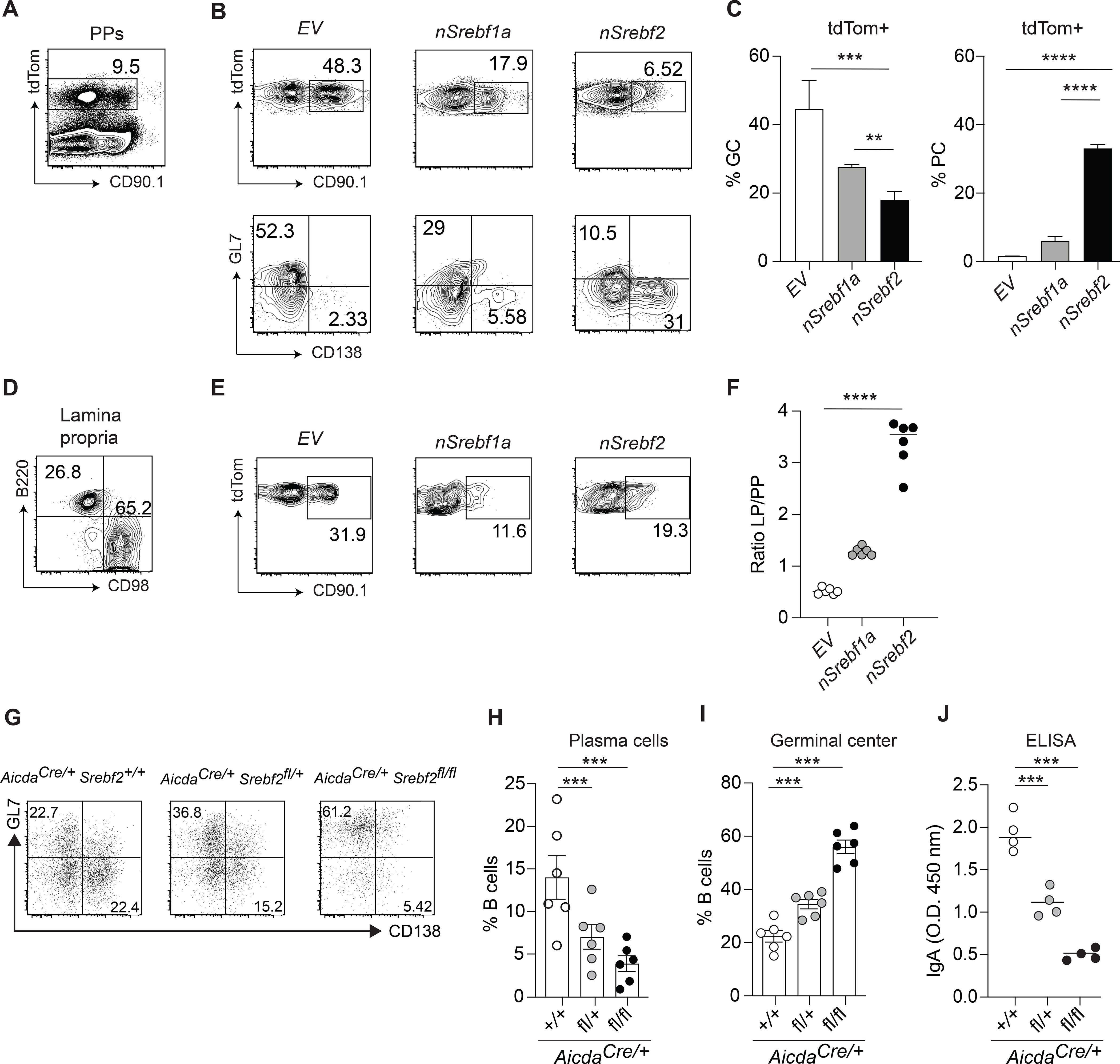
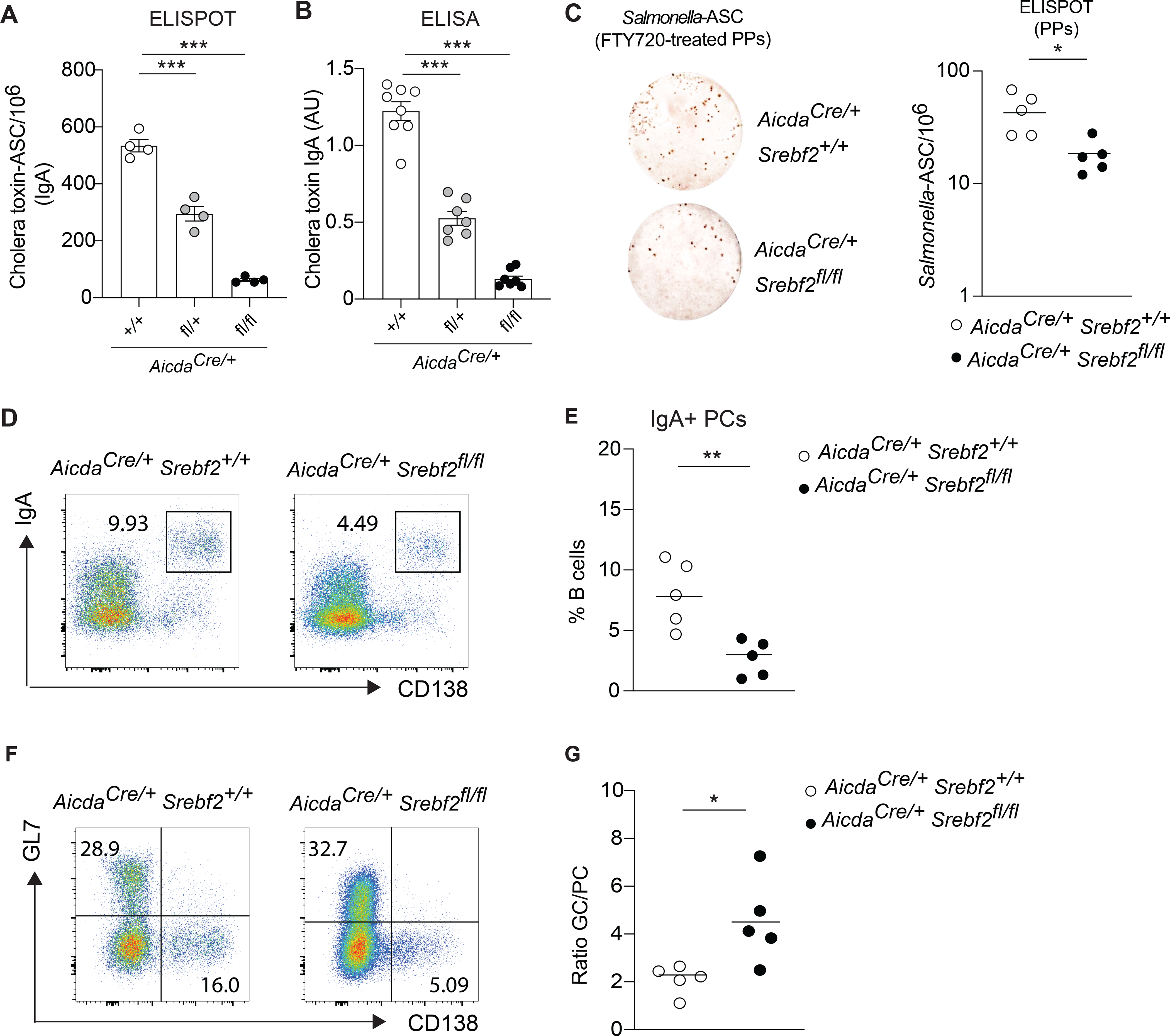
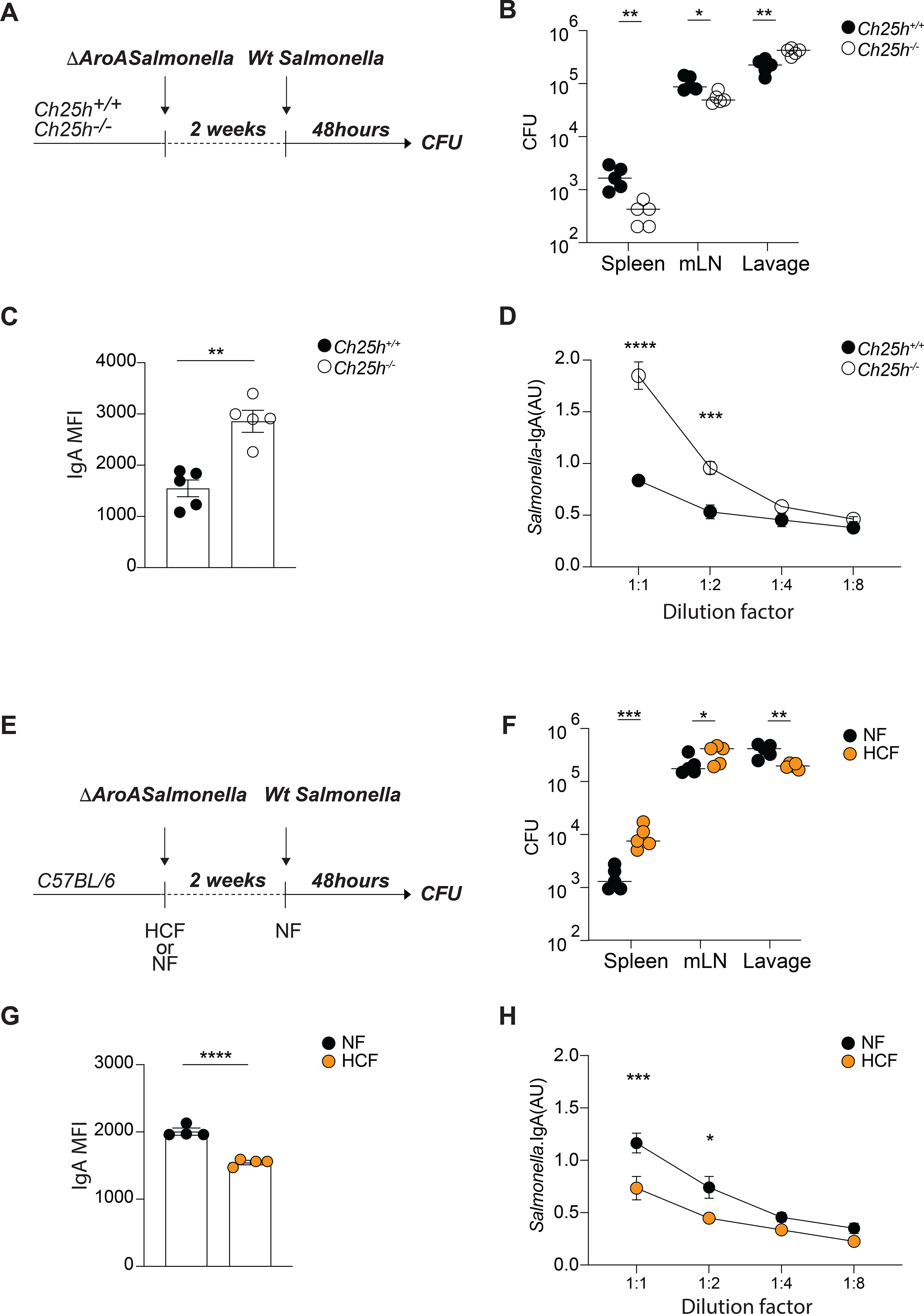
Comment in
-
25-hydroxycholesterol: Gatekeeper of intestinal IgA.Immunity. 2021 Oct 12;54(10):2182-2185. doi: 10.1016/j.immuni.2021.09.017. Immunity. 2021. PMID: 34644552
References
-
- Baptista AP, Gola A, Huang Y, Milanez-Almeida P, Torabi-Parizi P, Urban JF Jr., Shapiro VS, Gerner MY, Germain RN, 2019. The Chemoattractant Receptor Ebi2 Drives Intranodal Naive CD4+ T Cell Peripheralization to Promote Effective Adaptive Immunity. Immunity 50, 1188–1201.e6. doi: 10.1016/j.immuni.2019.04.001 - DOI - PubMed
-
- Blanc M, Hsieh WY, Robertson KA, Kropp KA, Forster T, Shui G, Lacaze P, Watterson S, Griffiths SJ, Spann NJ, Meljon A, Talbot S, Krishnan K, Covey DF, Wenk MR, Craigon M, Ruzsics Z, Haas J, Angulo A, Griffiths WJ, Glass CK, Wang Y, Ghazal P, 2012. The Transcription Factor STAT-1 Couples Macrophage Synthesis of 25-Hydroxycholesterol to the Interferon Antiviral Response. Immunity 1–13. doi: 10.1016/j.immuni.2012.11.004 - DOI - PMC - PubMed
-
- Buffie CG, Bucci V, Stein RR, McKenney PT, Ling L, Gobourne A, No D, Liu H, Kinnebrew M, Viale A, Littmann E, van den Brink MRM, Jenq RR, Taur Y, Sander C, Cross JR, Toussaint NC, Xavier JB, Pamer EG, 2014. Precision microbiome reconstitution restores bile acid mediated resistance to Clostridium difficile. Nature 517, 205–208. doi: 10.1038/nature13828 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
