

Astrocyte Ca2+-evoked ATP release regulates myelinated axon excitability and conduction speed
- PMID: 34648330
- PMCID: PMC7611967
- DOI: 10.1126/science.abh2858
Astrocyte Ca2+-evoked ATP release regulates myelinated axon excitability and conduction speed
Abstract
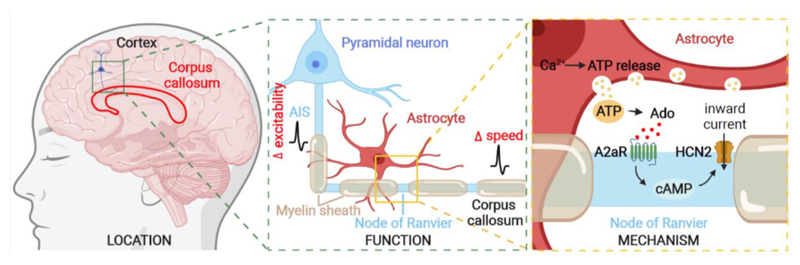
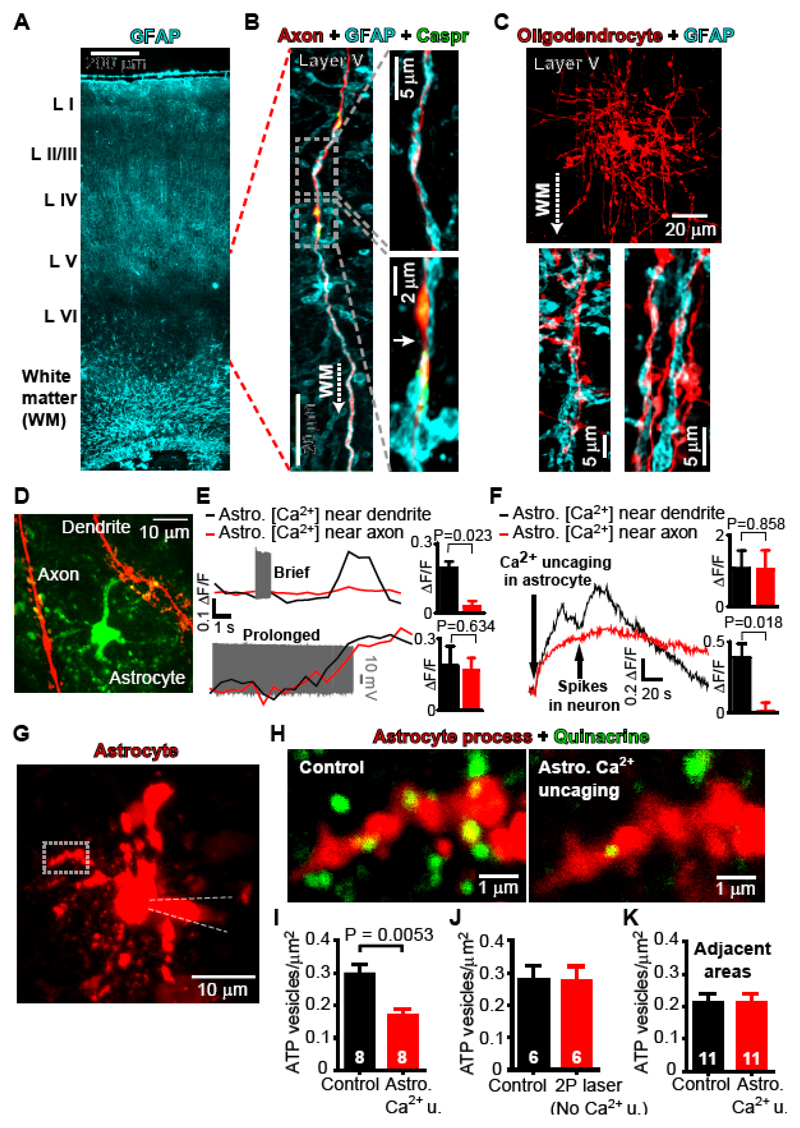
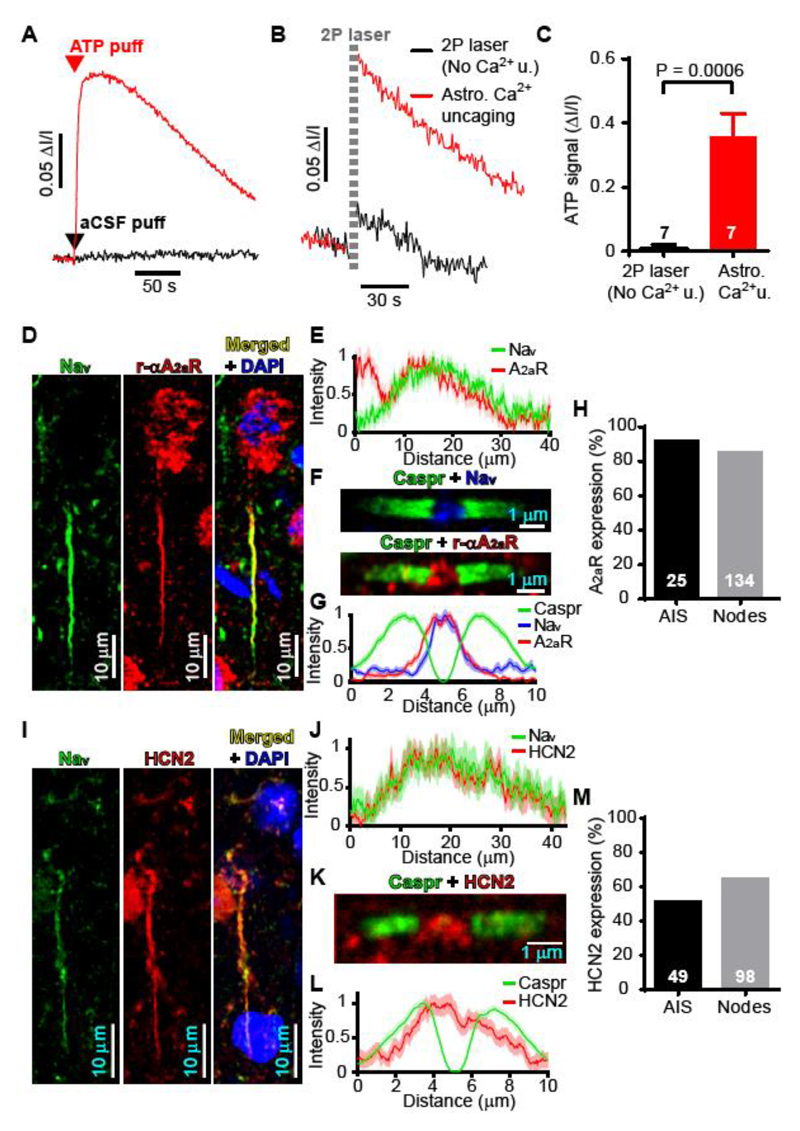
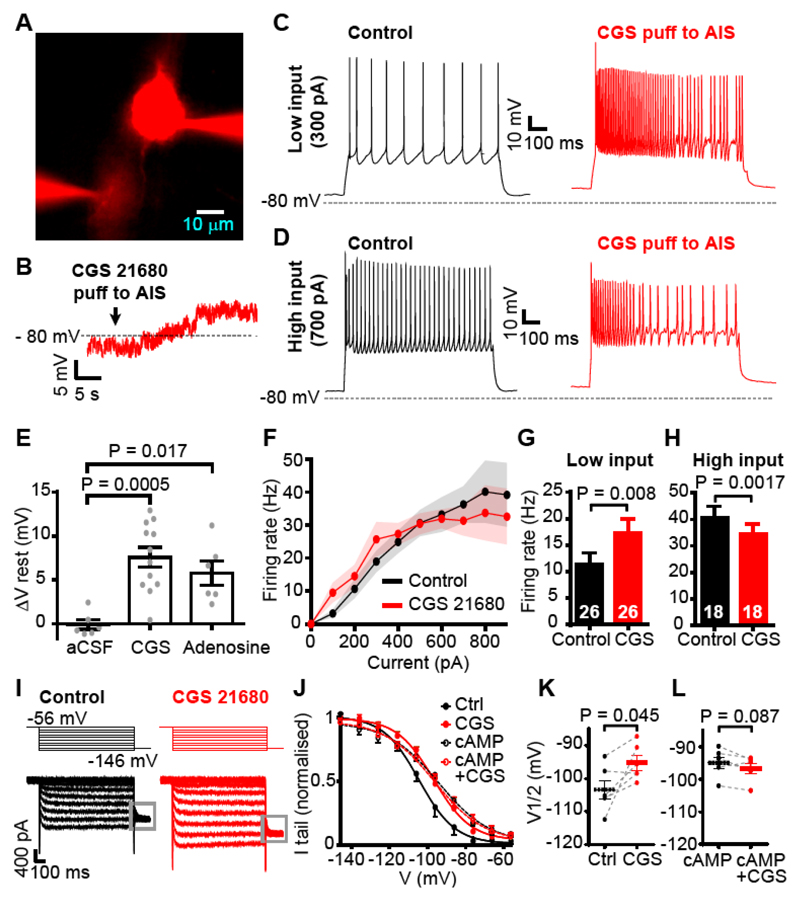
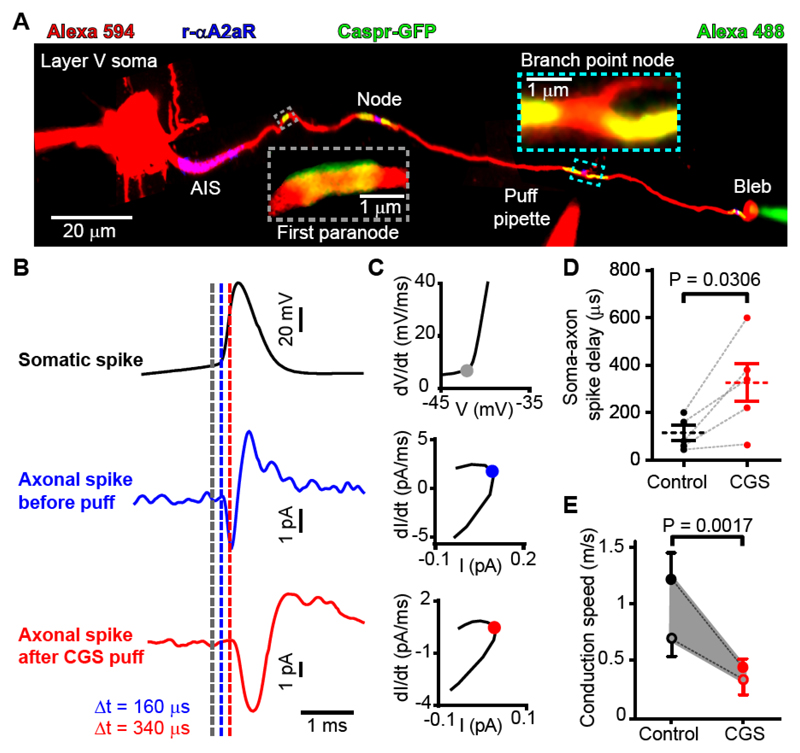
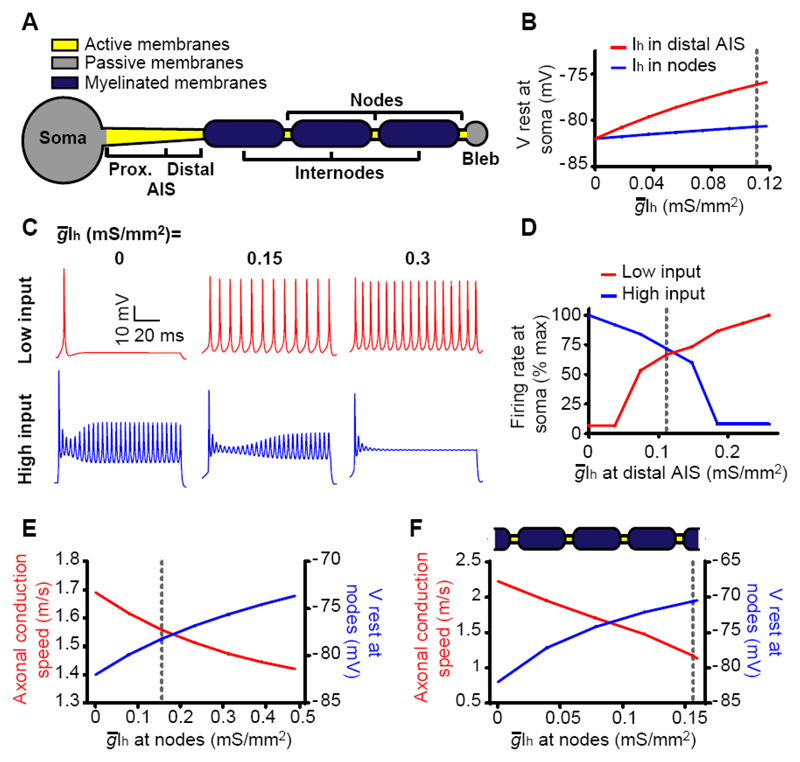
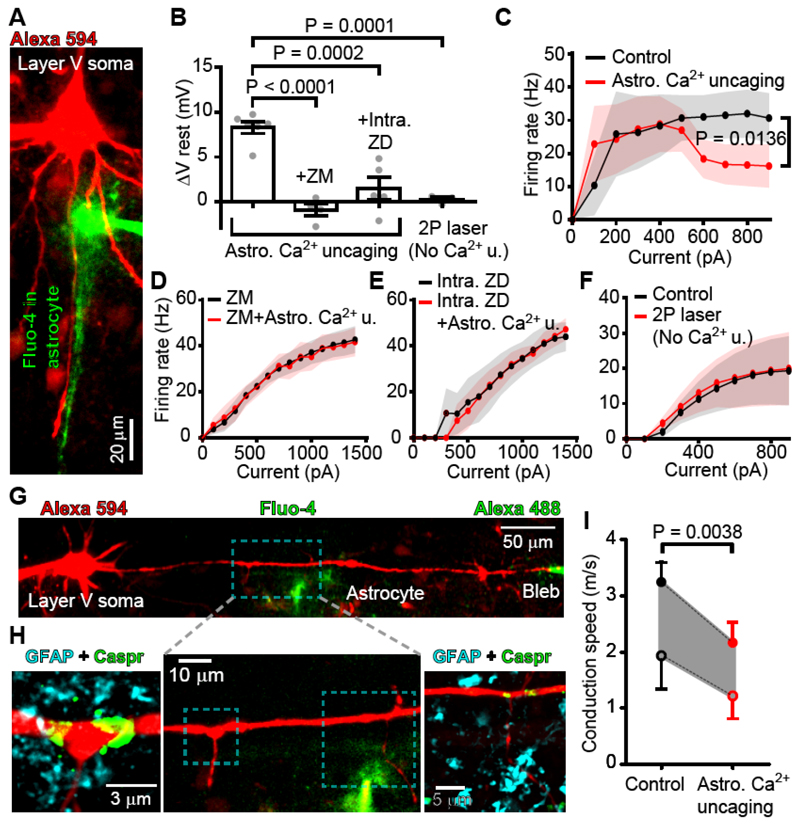
In the brain’s gray matter, astrocytes regulate synapse properties, but their role is unclear for the white matter, where myelinated axons rapidly transmit information between gray matter areas. We found that in rodents, neuronal activity raised the intracellular calcium concentration ([Ca2+]i) in astrocyte processes located near action potential–generating sites in the axon initial segment (AIS) and nodes of Ranvier of myelinated axons. This released adenosine triphosphate, which was converted extracellularly to adenosine and thus, through A2a receptors, activated HCN2-containing cation channels that regulate two aspects of myelinated axon function: excitability of the AIS and speed of action potential propagation. Variations in astrocyte-derived adenosine level between wake and sleep states or during energy deprivation could thus control white matter information flow and neural circuit function.
Conflict of interest statement
None.
Figures
References
-
- Allen NJ. Astrocyte regulation of synaptic behavior. Ann Rev Cell Dev Biol. 2014;30:439–463. - PubMed
-
- Kole MHP, Ilschner SU, Kampa BM, Williams SR, Ruben PC, Stuart GJ. Action potential generation requires a high sodium channel density in the axon initial segment. Nat Neurosci. 2008;11:178–186. - PubMed
-
- Saab AS, Tzvetanova ID, Nave K-A. The role of myelin and oligodendrocytes in axonal energy metabolism. Curr Opin Neurobiol. 2013;23:1065–1072. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
