

Redox Regulation in Cancer Cells during Metastasis
- PMID: 34649956
- PMCID: PMC8563381
- DOI: 10.1158/2159-8290.CD-21-0558
Redox Regulation in Cancer Cells during Metastasis
Abstract
Metastasis is an inefficient process in which the vast majority of cancer cells are fated to die, partly because they experience oxidative stress. Metastasizing cancer cells migrate through diverse environments that differ dramatically from their tumor of origin, leading to redox imbalances. The rare metastasizing cells that survive undergo reversible metabolic changes that confer oxidative stress resistance. We review the changes in redox regulation that cancer cells undergo during metastasis. By better understanding these mechanisms, it may be possible to develop pro-oxidant therapies that block disease progression by exacerbating oxidative stress in cancer cells. SIGNIFICANCE: Oxidative stress often limits cancer cell survival during metastasis, raising the possibility of inhibiting cancer progression with pro-oxidant therapies. This is the opposite strategy of treating patients with antioxidants, an approach that worsened outcomes in large clinical trials.
©2021 The Authors; Published by the American Association for Cancer Research.
Figures
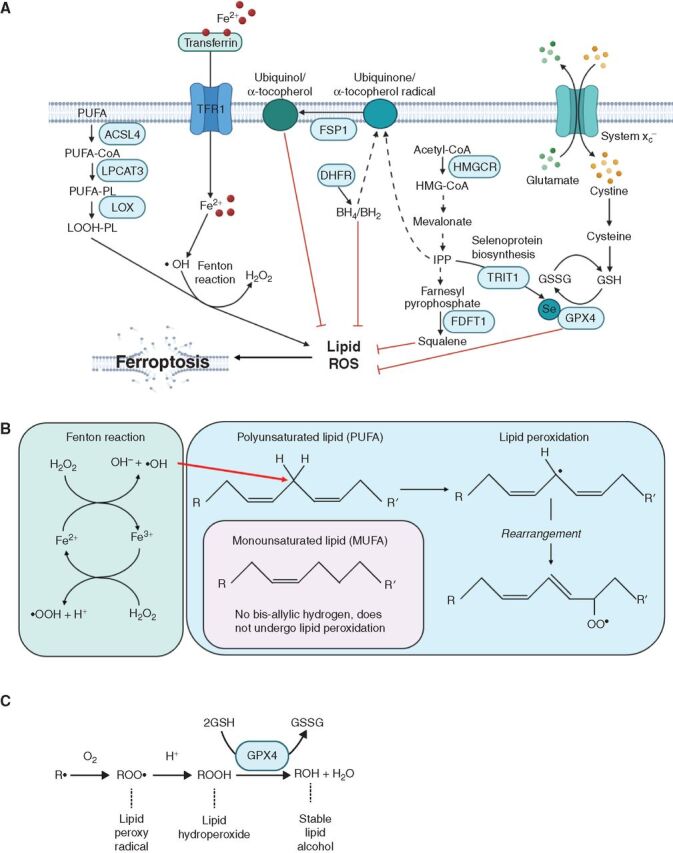
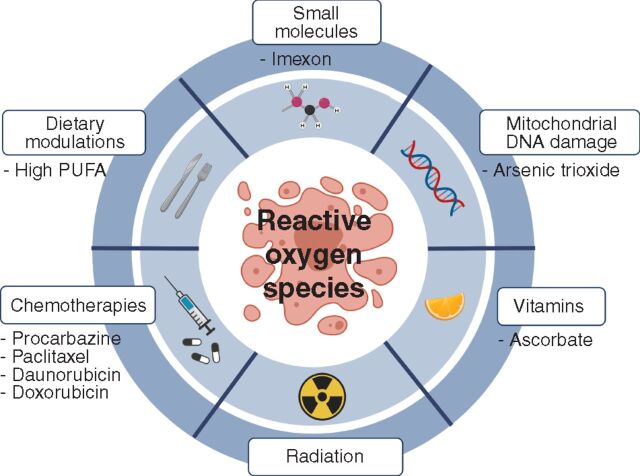
![Figure 1. Metabolic pathways that generate NADPH are important sources of reducing equivalents for oxidative stress resistance. A, Glutathione (GSH) and thioredoxin (TRXred) are redox buffers that are used by antioxidant enzymes such as superoxide dismutase (SOD), peroxiredoxin (PRDX), and glutathione peroxidase 4 (GPX4) to neutralize ROS, including O2−, H2O2, and lipid ROS. The reduced forms of GSH and TRX can then be regenerated from the oxidized forms [glutathione disulfide (GSSG); TRXox] by glutathione reductase (GR) or thioredoxin reductase (TRXR), which obtain reducing equivalents from NADPH. NADP+ is generated de novo from NAD+ by NAD kinase (NADK; ref. 167). NADP+ is then reduced to NADPH by the pentose phosphate pathway, the folate pathway, malic enzyme (ME1, 2, or 3), glutamate dehydrogenase (GDH1/2), or isocitrate dehydrogenase (IDH1/2; ref. 86). Other abbreviations in this panel include electron transport chain (ETC), glucose-6-phosphate dehydrogenase (G6PD), phosphogluconate dehydrogenase (PGD), dihydrofolate reductase (DHFR), methylenetetrahydrofolate dehydrogenase 1/2 (MTHFD1/2), NADPH oxidase (NOX), superoxide dismutase (SOD), and catalase (CAT). B, Schematic of reactions in which antioxidant enzymes transfer reducing equivalents between NADPH, GSH, and ROS.](https://cdn.ncbi.nlm.nih.gov/pmc/blobs/79cb/9306307/c469072f6481/2682fig1.jpg)
References
-
- Cameron MD, Schmidt EE, Kerkvliet N, Nadkarni KV, Morris VL, Groom AC, et al. Temporal progression of metastasis in lung: cell survival, dormancy, and location dependence of metastatic inefficiency. Cancer Res 2000;60:2541–6. - PubMed
-
- Kienast Y, von Baumgarten L, Fuhrmann M, Klinkert WE, Goldbrunner R, Herms J, et al. Real-time imaging reveals the single steps of brain metastasis formation. Nat Med 2010;16:116–22. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
