

Precise genomic deletions using paired prime editing
- PMID: 34650269
- PMCID: PMC8847327
- DOI: 10.1038/s41587-021-01025-z
Precise genomic deletions using paired prime editing
Abstract
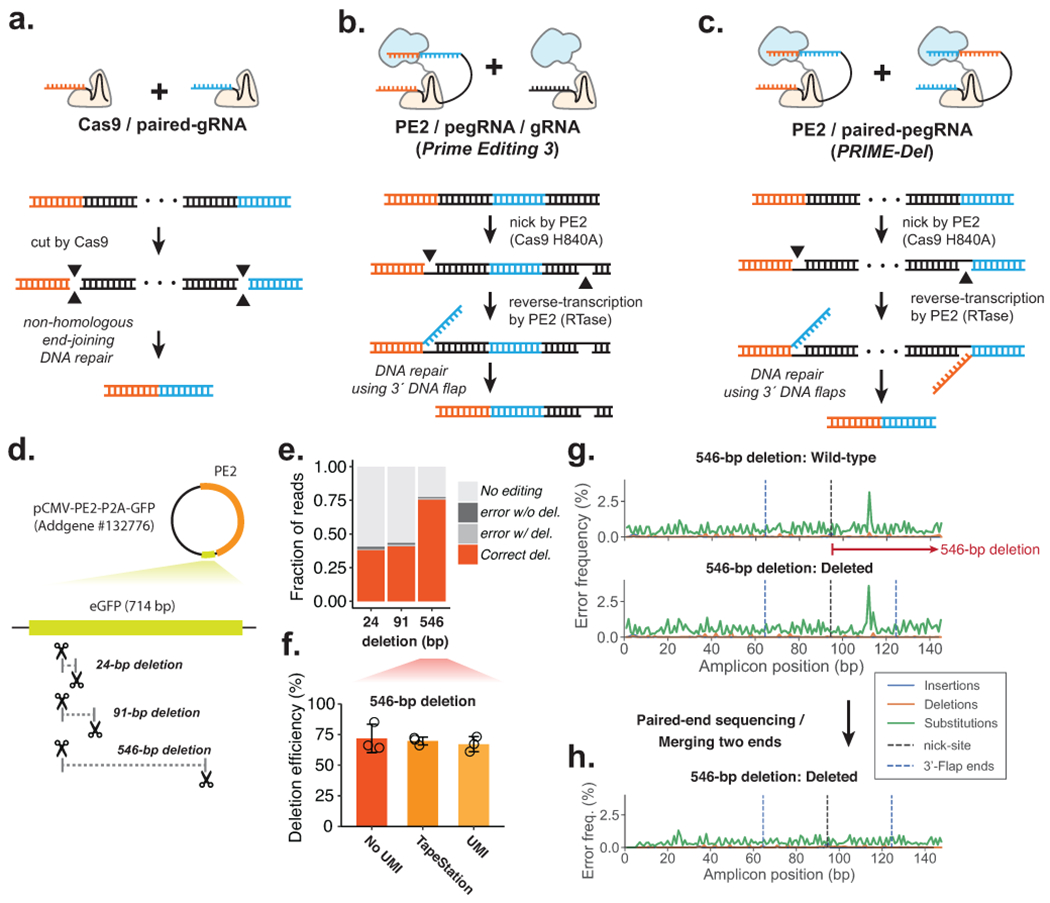
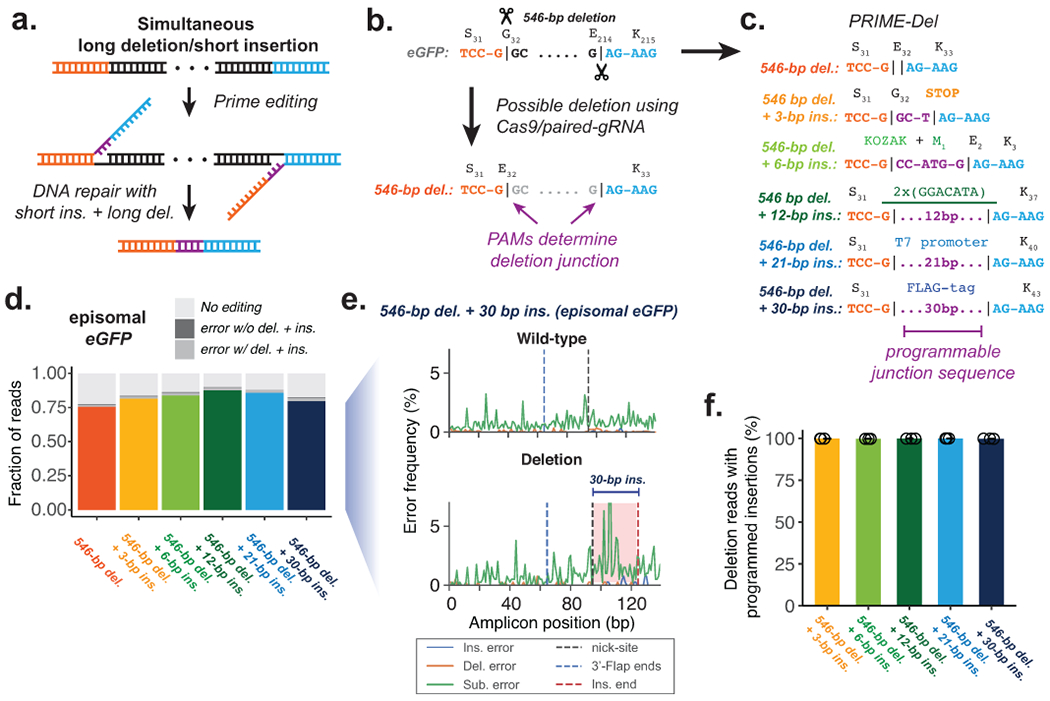
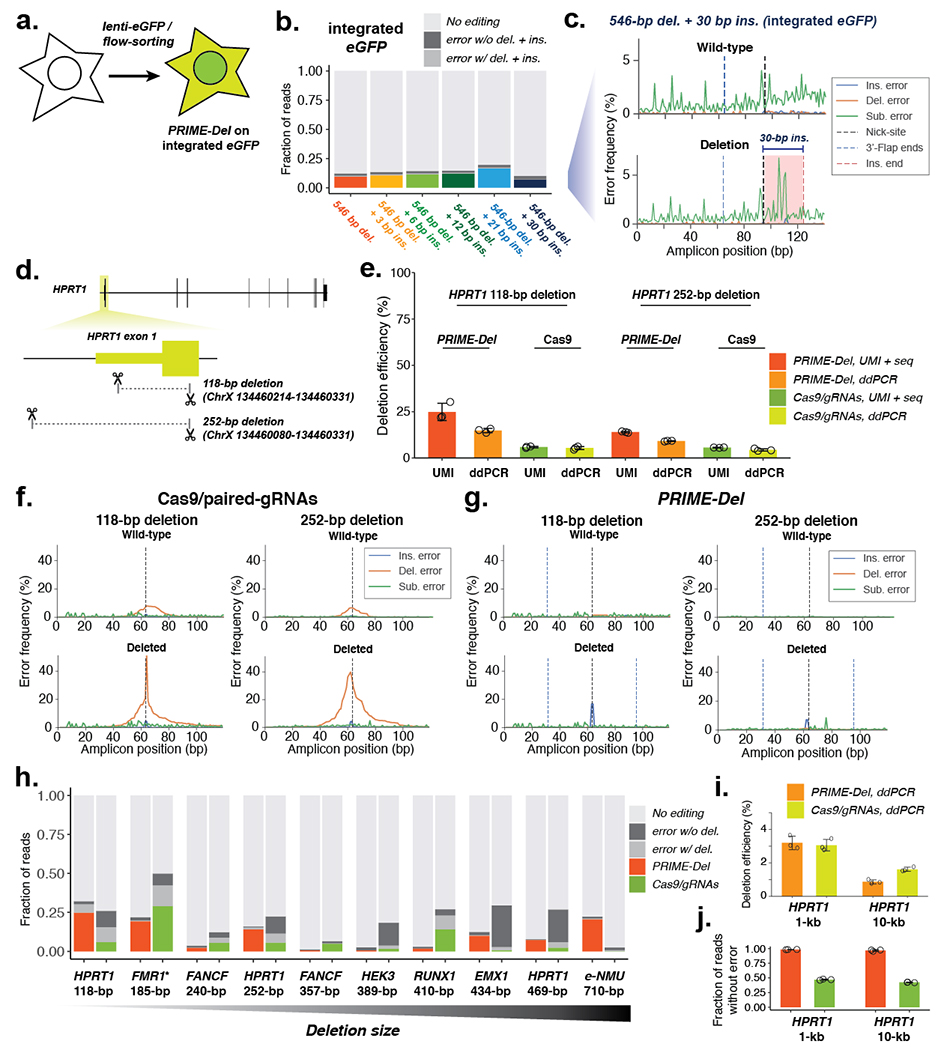
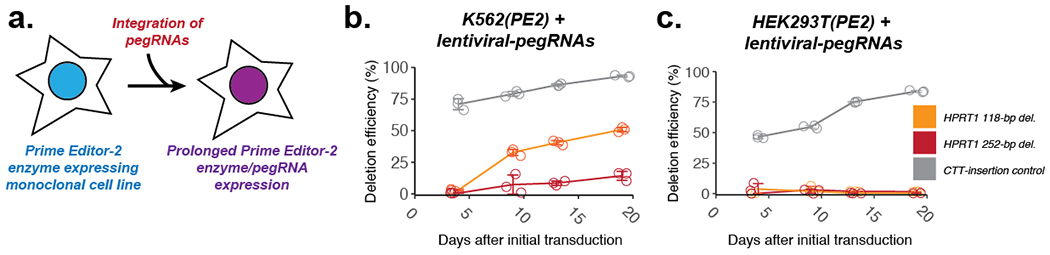
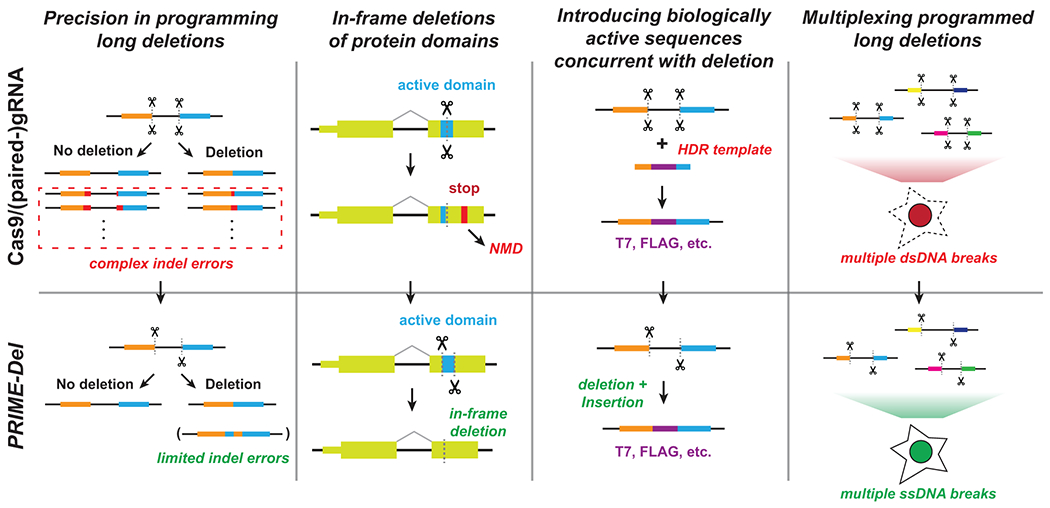
Current methods to delete genomic sequences are based on clustered regularly interspaced short palindromic repeats (CRISPR)-Cas9 and pairs of single-guide RNAs (sgRNAs), but can be inefficient and imprecise, with errors including small indels as well as unintended large deletions and more complex rearrangements. In the present study, we describe a prime editing-based method, PRIME-Del, which induces a deletion using a pair of prime editing sgRNAs (pegRNAs) that target opposite DNA strands, programming not only the sites that are nicked but also the outcome of the repair. PRIME-Del achieves markedly higher precision than CRISPR-Cas9 and sgRNA pairs in programming deletions up to 10 kb, with 1-30% editing efficiency. PRIME-Del can also be used to couple genomic deletions with short insertions, enabling deletions with junctions that do not fall at protospacer-adjacent motif sites. Finally, extended expression of prime editing components can substantially enhance efficiency without compromising precision. We anticipate that PRIME-Del will be broadly useful for precise, flexible programming of genomic deletions, epitope tagging and, potentially, programming genomic rearrangements.
© 2021. The Author(s), under exclusive licence to Springer Nature America, Inc.
Conflict of interest statement
Competing interests
The University of Washington has filed a patent application based on this work, in which J.C., W.C., and J.S. are listed as inventors.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
