

Sepsis leads to lasting changes in phenotype and function of memory CD8 T cells
- PMID: 34652273
- PMCID: PMC8589447
- DOI: 10.7554/eLife.70989
Sepsis leads to lasting changes in phenotype and function of memory CD8 T cells
Abstract
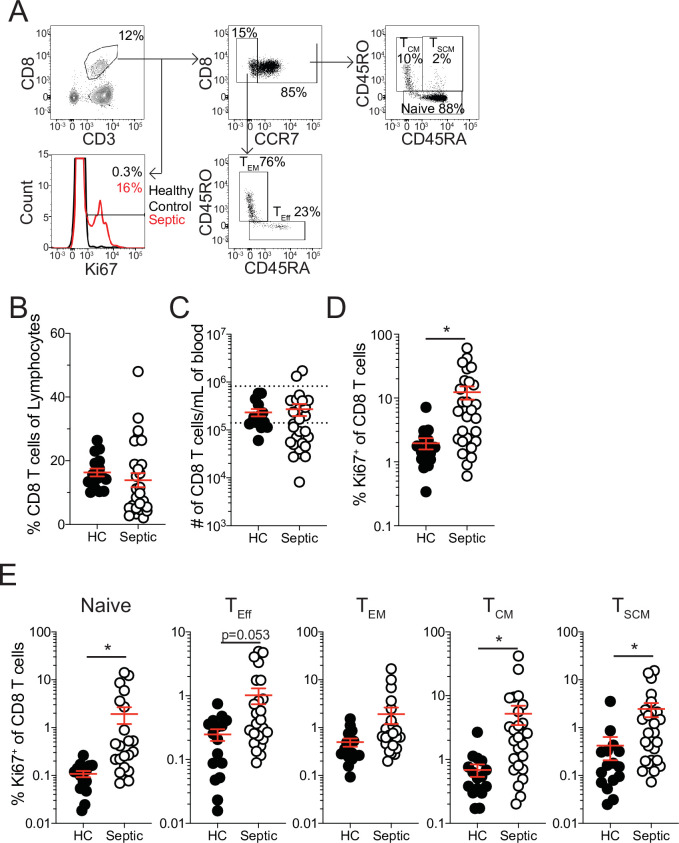
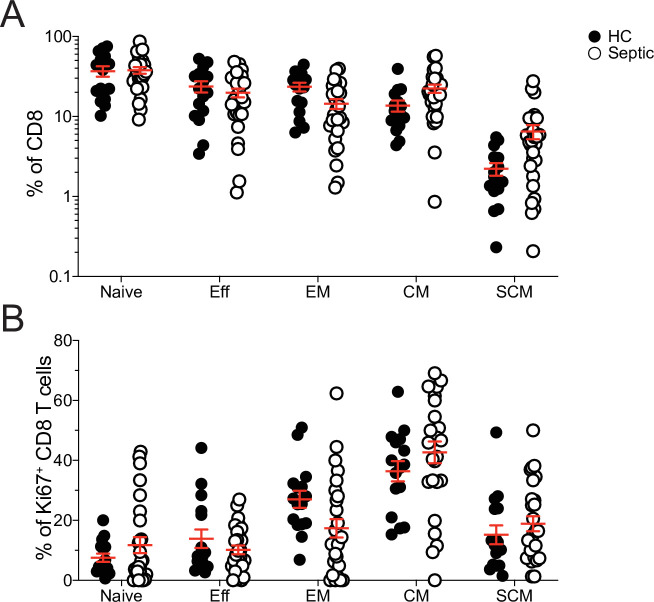
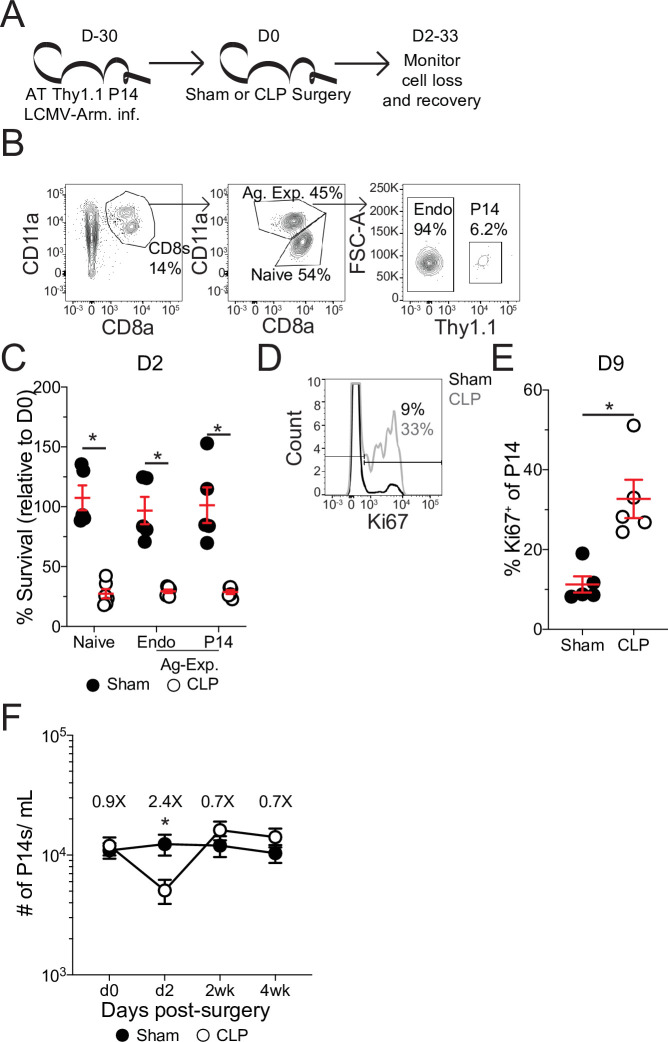
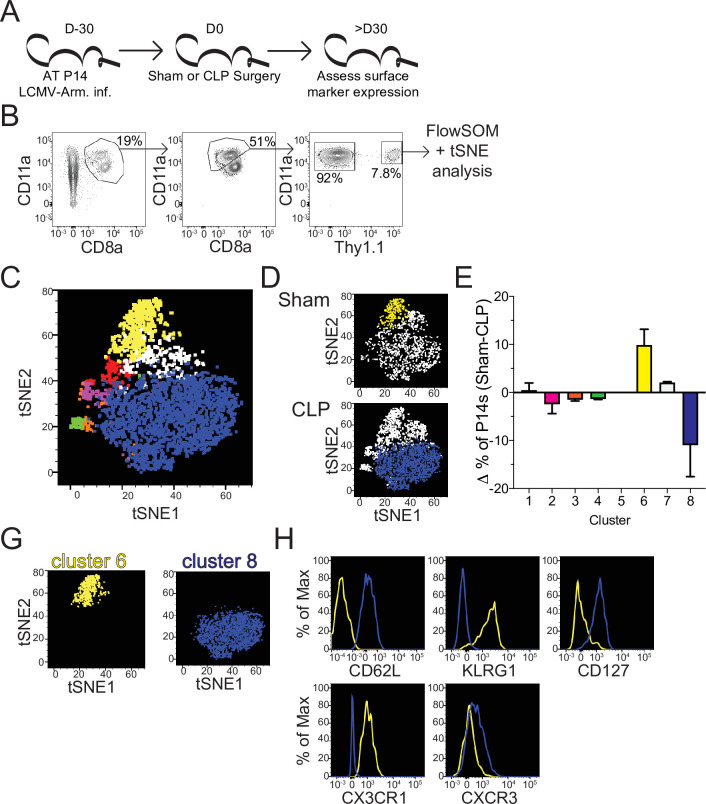
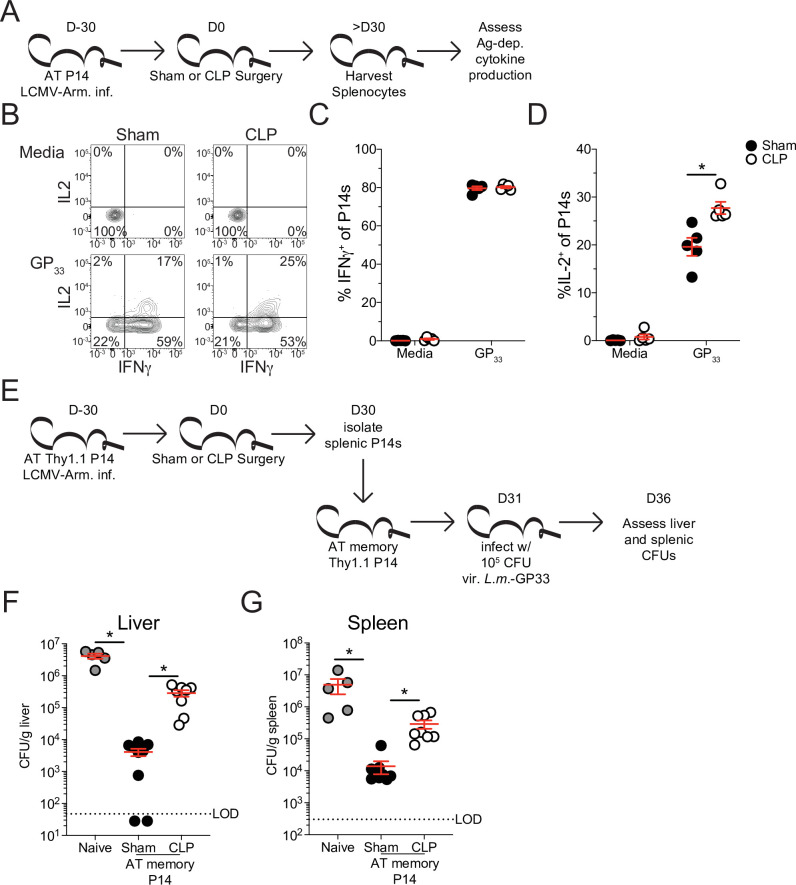
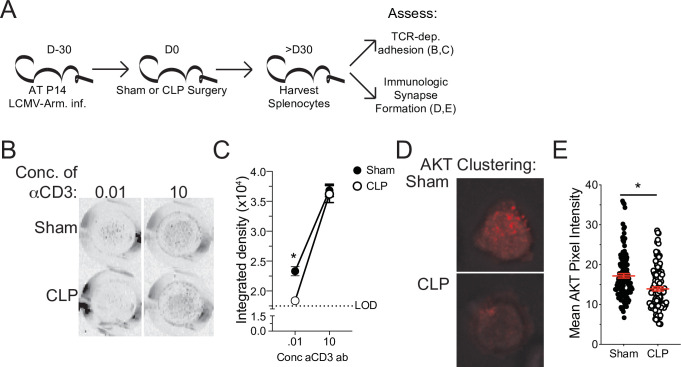
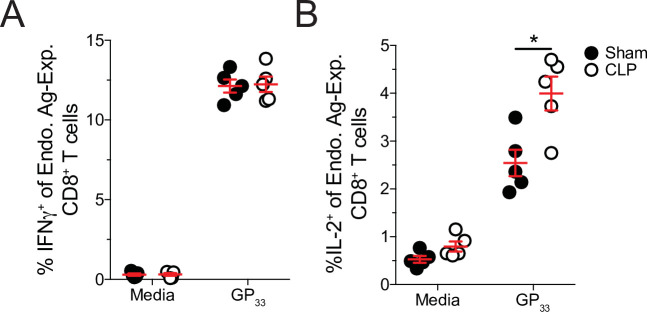
The global health burden due to sepsis and the associated cytokine storm is substantial. While early intervention has improved survival during the cytokine storm, those that survive can enter a state of chronic immunoparalysis defined by transient lymphopenia and functional deficits of surviving cells. Memory CD8 T cells provide rapid cytolysis and cytokine production following re-encounter with their cognate antigen to promote long-term immunity, and CD8 T cell impairment due to sepsis can pre-dispose individuals to re-infection. While the acute influence of sepsis on memory CD8 T cells has been characterized, if and to what extent pre-existing memory CD8 T cells recover remains unknown. Here, we observed that central memory CD8 T cells (TCM) from septic patients proliferate more than those from healthy individuals. Utilizing LCMV immune mice and a CLP model to induce sepsis, we demonstrated that TCM proliferation is associated with numerical recovery of pathogen-specific memory CD8 T cells following sepsis-induced lymphopenia. This increased proliferation leads to changes in composition of memory CD8 T cell compartment and altered tissue localization. Further, memory CD8 T cells from sepsis survivors have an altered transcriptional profile and chromatin accessibility indicating long-lasting T cell intrinsic changes. The sepsis-induced changes in the composition of the memory CD8 T cell pool and transcriptional landscape culminated in altered T cell function and reduced capacity to control L. monocytogenes infection. Thus, sepsis leads to long-term alterations in memory CD8 T cell phenotype, protective function and localization potentially changing host capacity to respond to re-infection.
Keywords: CD8 T cell; central memory; homeostatic proliferation; human; immunology; inflammation; mouse; sepsis.
Plain language summary
A dirty cut, a nasty burn, a severe COVID infection; there are many ways for someone to develop sepsis. This life-threatening condition emerges when the immune system overreacts to a threat and ends up damaging the body. Even when patients survive, they are often left with a partially impaired immune system that cannot adequately protect against microbes and cancer; this is known as immunoparalysis. Memory CD8 T cells, a type of immune cell that is compromised by sepsis, are a long-lived population of cells that ‘remember’ previous infection or vaccination, and then react faster to prevent the same illness if the person ever encounters the same threat again. Yet it is unclear how exactly sepsis harms the function and representation of memory CD8 T cells, and the immune system in general. Jensen et al. investigated this question, first by showing that sepsis leads to a profound loss of memory CD8 T cells, but that surviving memory CD8 T cells multiply quickly – especially a subpopulation known as central memory CD8 T cells – to re-establish the memory CD8 T cell population. Since the central memory CD8 T cells proliferate better than the other memory T cells this alters the overall composition of the pool of memory CD8 T cells, with central memory cells becoming overrepresented. Further experiments revealed that this biasing toward central memory T cells, due to sepsis, created long-term changes in the distribution of memory CD8 T cells throughout the body. The way the genetic information of these cells was packaged had also been altered, as well as which genes were switched on or off. Overall, these changes reduced the ability of memory CD8 T cells to control infections. Together, these findings help to understand how immunoparalysis can emerge after sepsis, and what could be done to correct it. These findings could also be applied to other conditions – such as COVID-19 – which may cause similar long-term changes to the immune system.
© 2021, Jensen et al.
Conflict of interest statement
IJ, XL, PM, QS, MF, MT, JH, HX, TG, WP, VB No competing interests declared
Figures
References
-
- Baek KH, Shin HJ, Yoo JK, Cho JH, Choi YH, Sung YC, McKeon F, Lee CW. p53 deficiency and defective mitotic checkpoint in proliferating T lymphocytes increase chromosomal instability through aberrant exit from mitotic arrest. Journal of Leukocyte Biology. 2003;73:850–861. doi: 10.1189/jlb.1202607. - DOI - PubMed
-
- Borges da Silva H, Beura LK, Wang H, Hanse EA, Gore R, Scott MC, Walsh DA, Block KE, Fonseca R, Yan Y, Hippen KL, Blazar BR, Masopust D, Kelekar A, Vulchanova L, Hogquist KA, Jameson SC. The purinergic receptor P2RX7 directs metabolic fitness of long-lived memory CD8+ T cells. Nature. 2018;559:264–268. doi: 10.1038/s41586-018-0282-0. - DOI - PMC - PubMed
Publication types
MeSH terms
Associated data
- Actions
Grants and funding
- R35 GM134880/GM/NIGMS NIH HHS/United States
- R35GM134880/NH/NIH HHS/United States
- R01 AI139874/AI/NIAID NIH HHS/United States
- R01AI121080/NH/NIH HHS/United States
- P30 CA086862/CA/NCI NIH HHS/United States
- T32AI007511/NH/NIH HHS/United States
- R01GM115462/NH/NIH HHS/United States
- P30CA086862/CA/NCI NIH HHS/United States
- R01 AI112579/AI/NIAID NIH HHS/United States
- R01AI139874/NH/NIH HHS/United States
- I0BX001324/Veterans Health Administration
- T32 AI007511/AI/NIAID NIH HHS/United States
- R21 AI151183/AI/NIAID NIH HHS/United States
- T32AI007485/NH/NIH HHS/United States
- R35GM140881/NH/NIH HHS/United States
- R01AI112579/NH/NIH HHS/United States
- T32 AI007485/AI/NIAID NIH HHS/United States
- I01 BX001324/BX/BLRD VA/United States
- I01 BX000207/BX/BLRD VA/United States
- R21AI147064/NH/NIH HHS/United States
- R01 GM115462/GM/NIGMS NIH HHS/United States
- R21 AI147064/AI/NIAID NIH HHS/United States
- R01 AI114543/AI/NIAID NIH HHS/United States
- T32 AI007343/AI/NIAID NIH HHS/United States
- R21AI157121/NH/NIH HHS/United States
- R01 AI121080/AI/NIAID NIH HHS/United States
- R21 AI157121/AI/NIAID NIH HHS/United States
- R01AI114543/NH/NIH HHS/United States
- R21AI151183/NH/NIH HHS/United States
- I0BX001324/Veterans Health Administration HSR and D
- R35 GM140881/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
