Functional attractors in microbial community assembly
- PMID: 34653368
- PMCID: PMC8800145
- DOI: 10.1016/j.cels.2021.09.011
Functional attractors in microbial community assembly
Abstract
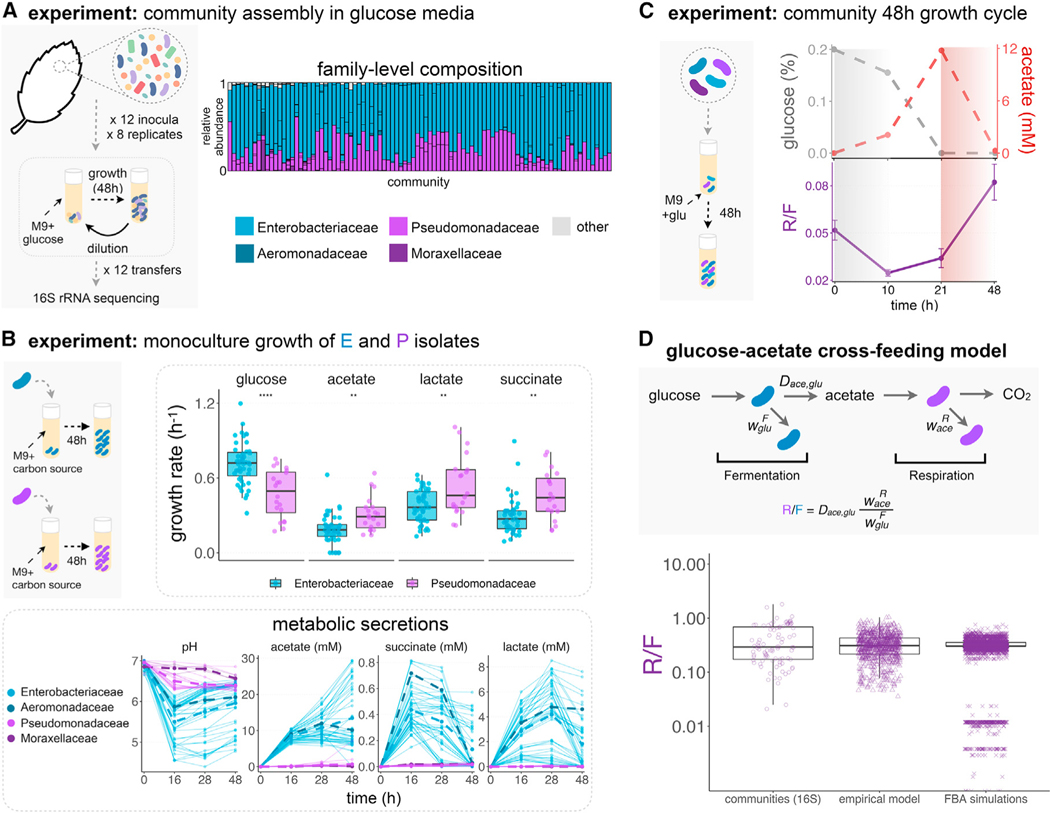
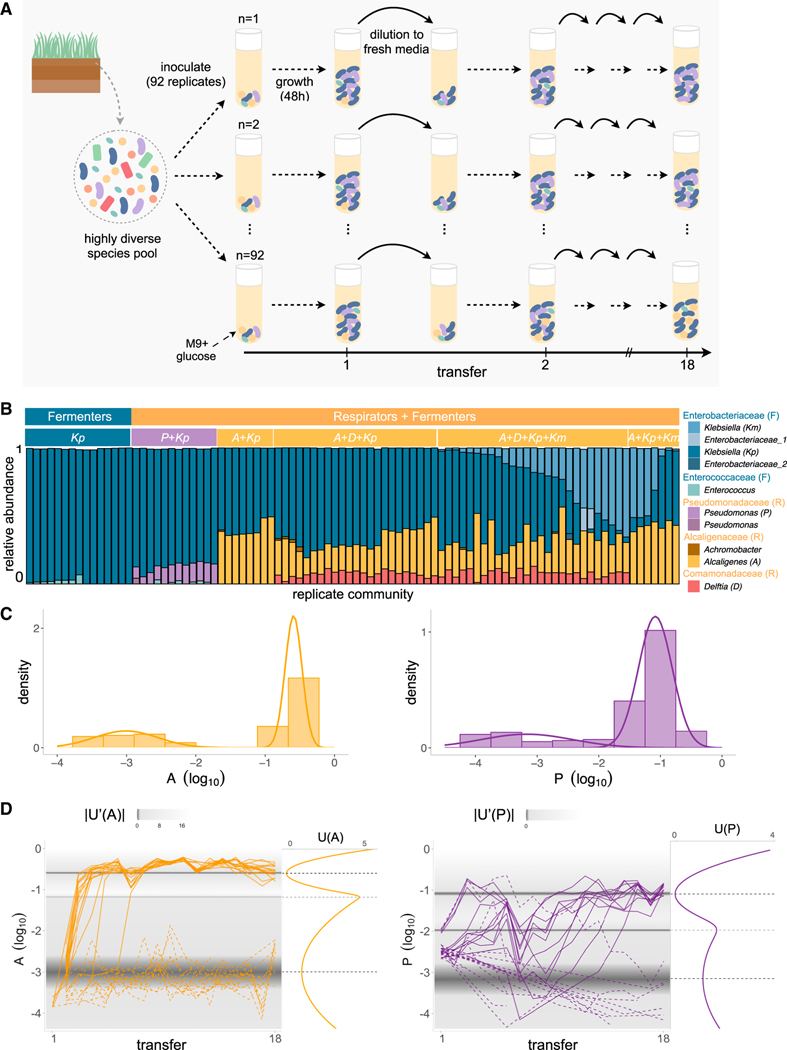
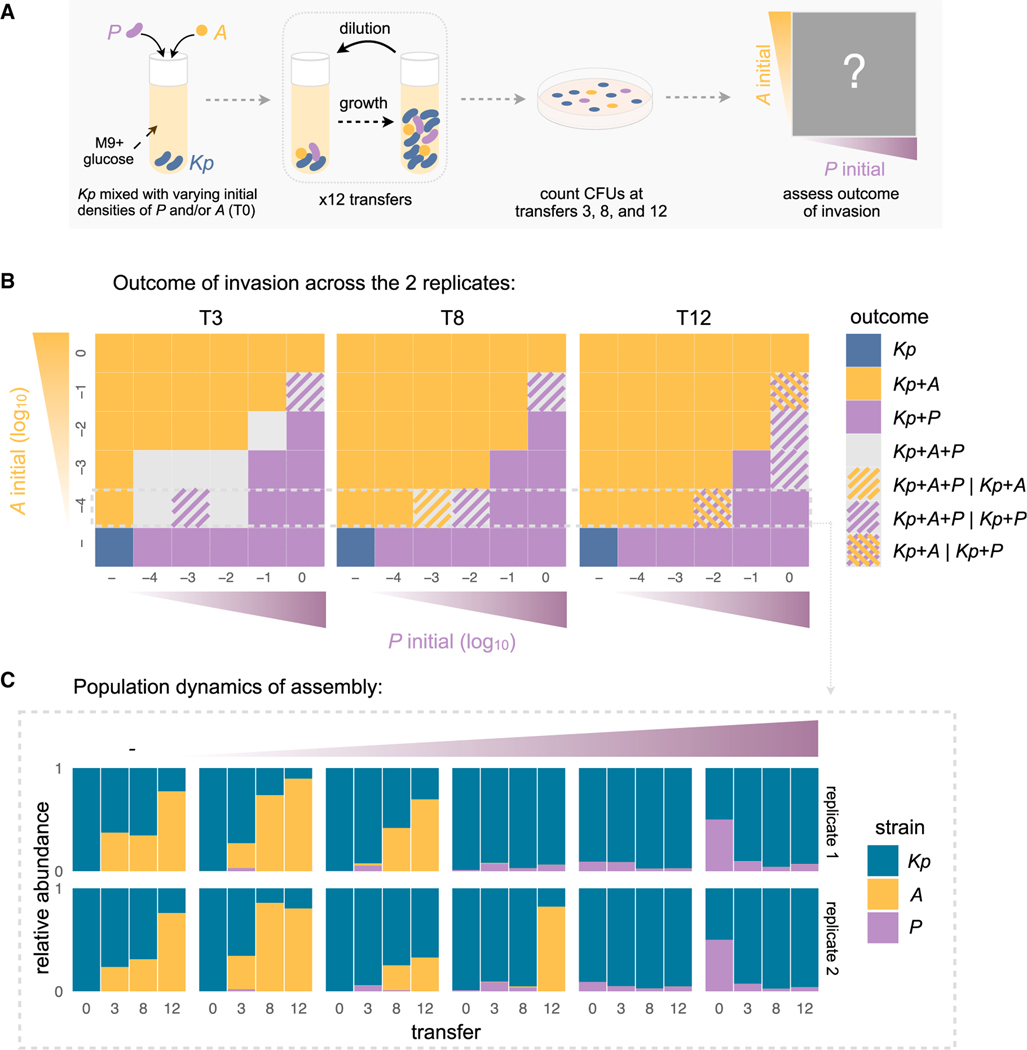
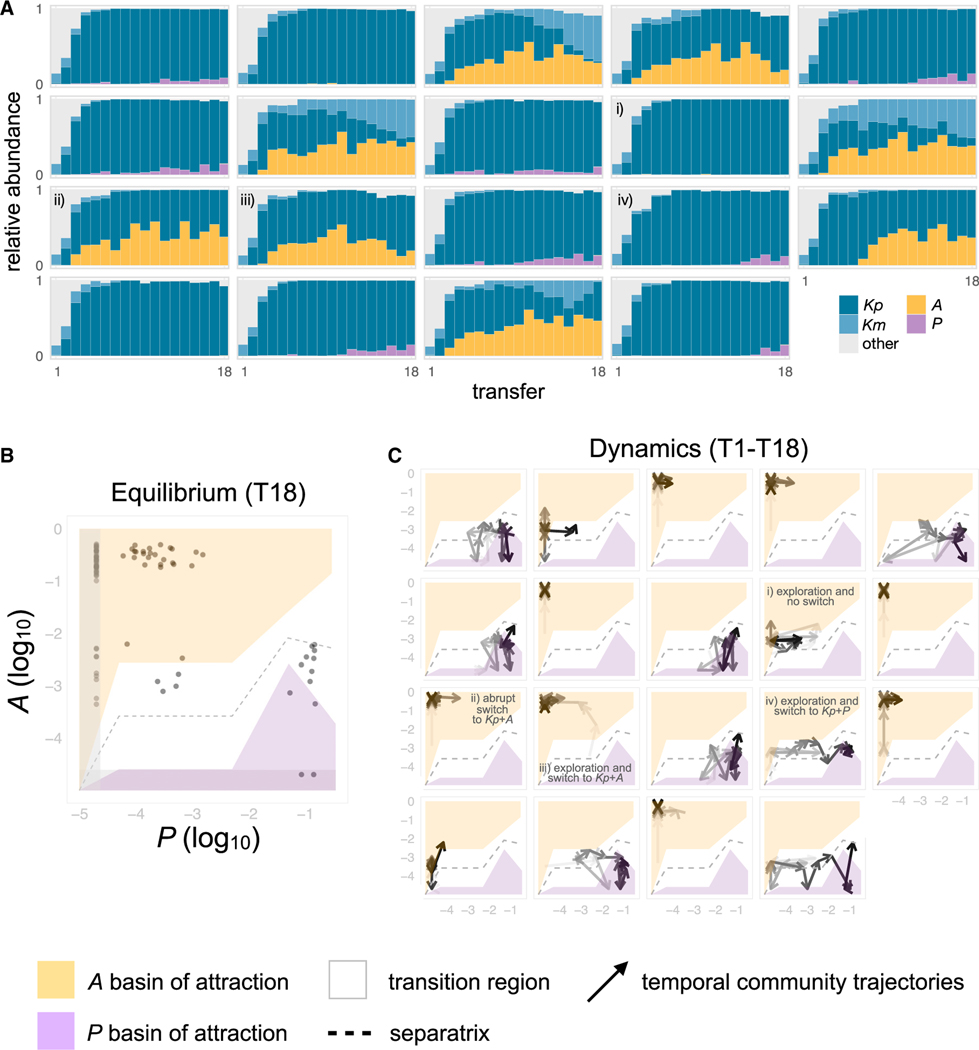
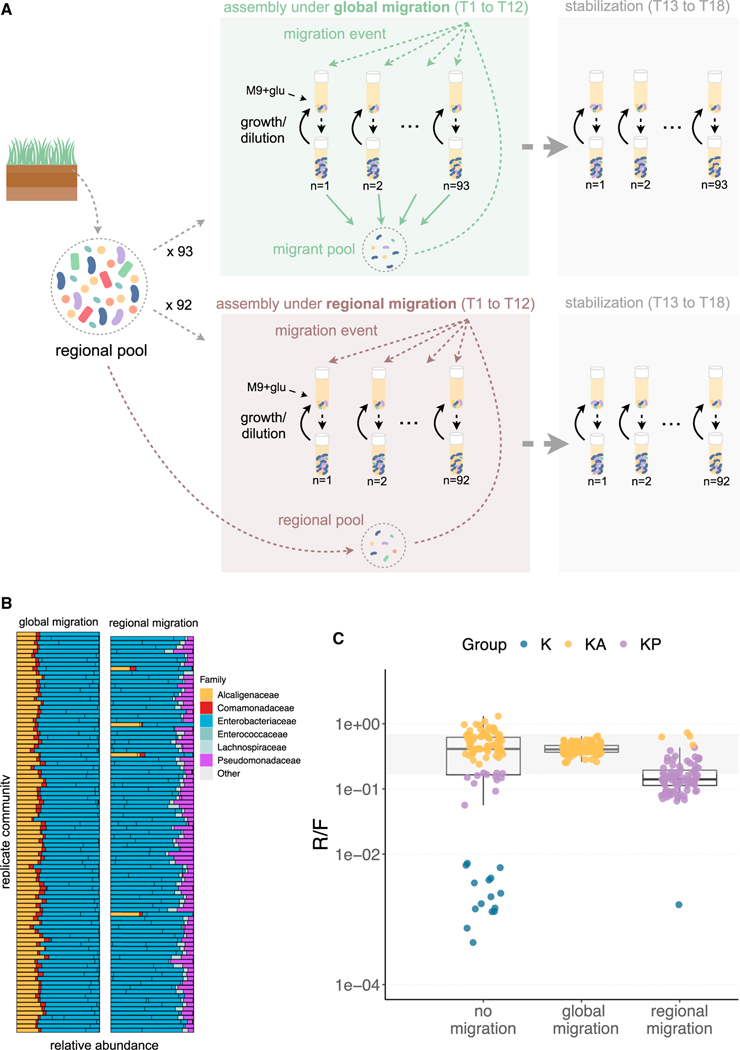
For microbiome biology to become a more predictive science, we must identify which descriptive features of microbial communities are reproducible and predictable, which are not, and why. We address this question by experimentally studying parallelism and convergence in microbial community assembly in replicate glucose-limited habitats. Here, we show that the previously observed family-level convergence in these habitats reflects a reproducible metabolic organization, where the ratio of the dominant metabolic groups can be explained from a simple resource-partitioning model. In turn, taxonomic divergence among replicate communities arises from multistability in population dynamics. Multistability can also lead to alternative functional states in closed ecosystems but not in metacommunities. Our findings empirically illustrate how the evolutionary conservation of quantitative metabolic traits, multistability, and the inherent stochasticity of population dynamics, may all conspire to generate the patterns of reproducibility and variability at different levels of organization that are commonplace in microbial community assembly.
Keywords: alternative states; dynamical systems theory; microbial community assembly; microbial metabolism; population dynamics.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests A.S. is a member of the Cell Systems Advisory Board. The authors otherwise declare no competing interests.
Figures