

Allele-specific gene editing to rescue dominant CRX-associated LCA7 phenotypes in a retinal organoid model
- PMID: 34653402
- PMCID: PMC8580887
- DOI: 10.1016/j.stemcr.2021.09.007
Allele-specific gene editing to rescue dominant CRX-associated LCA7 phenotypes in a retinal organoid model
Abstract
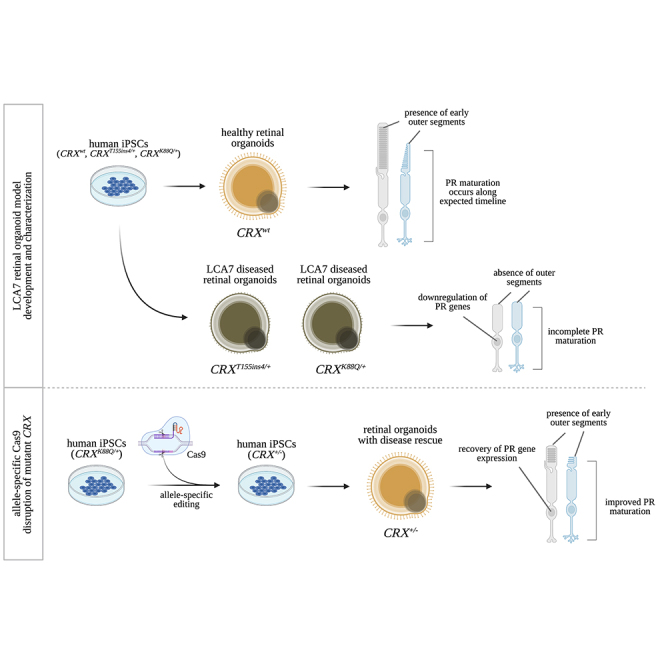
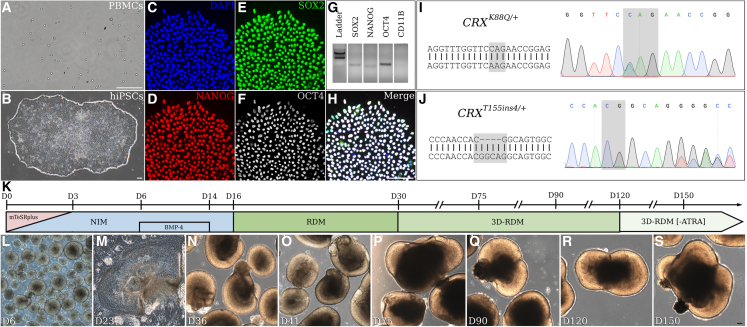
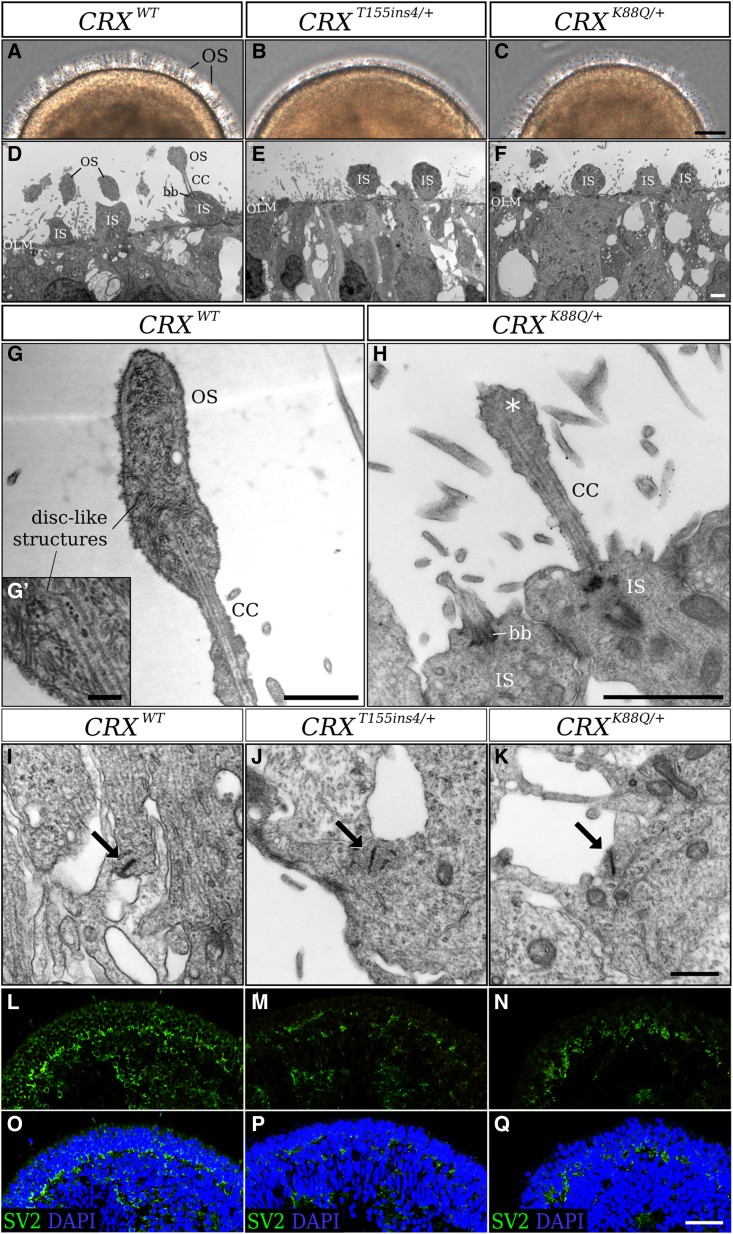
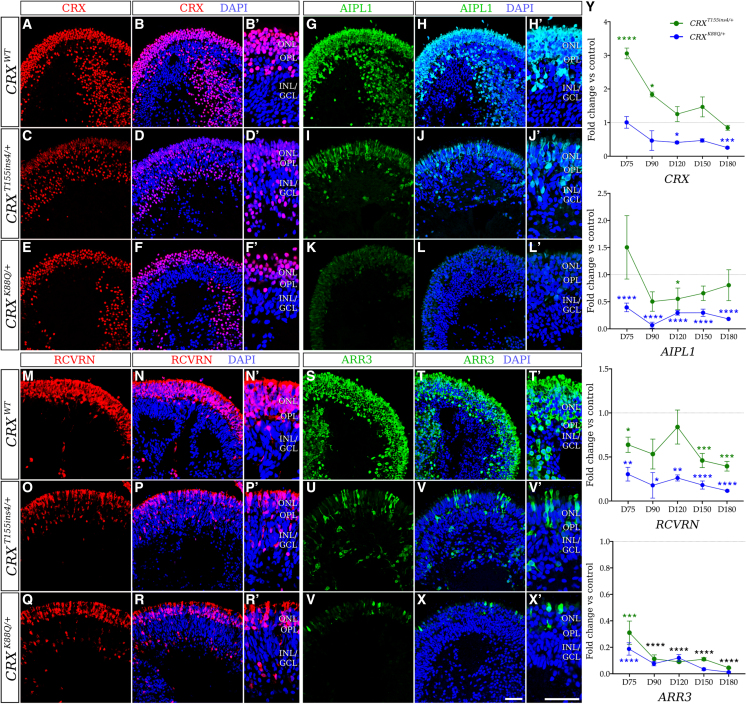
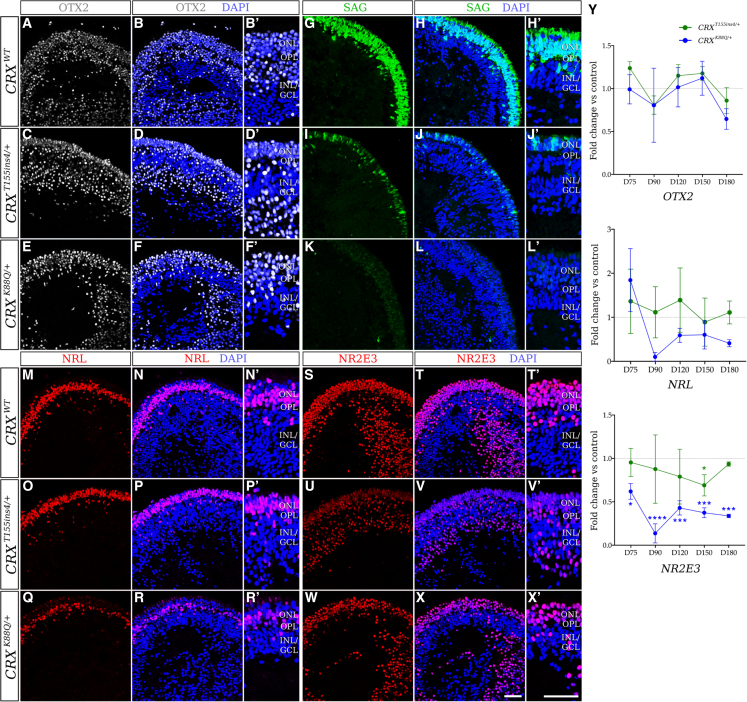
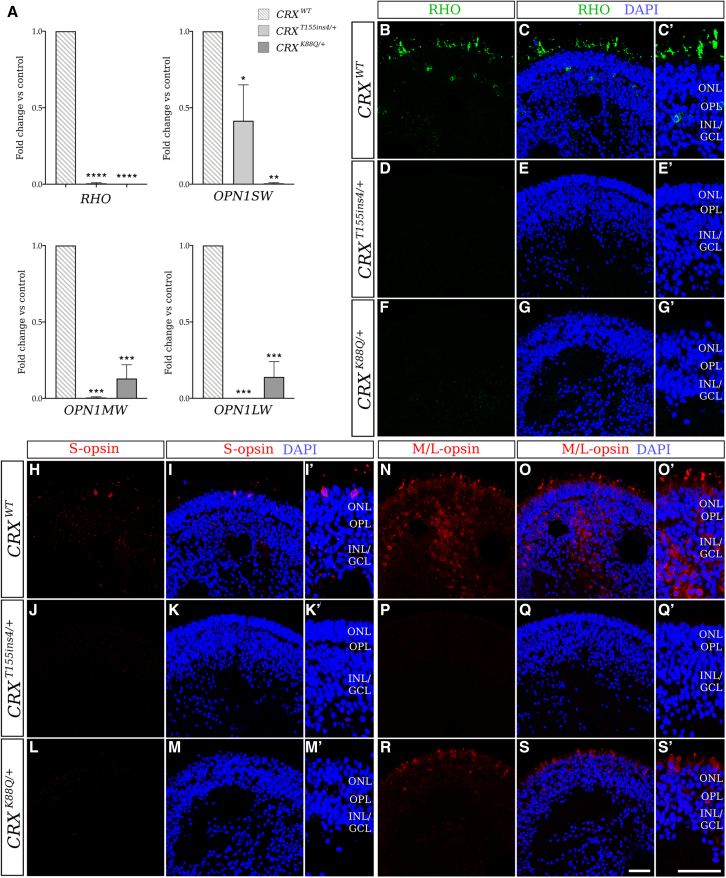
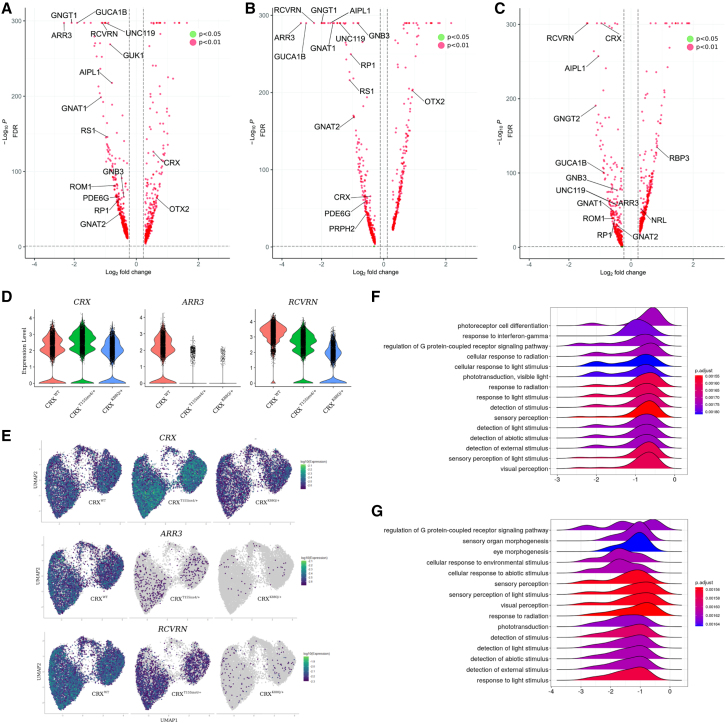
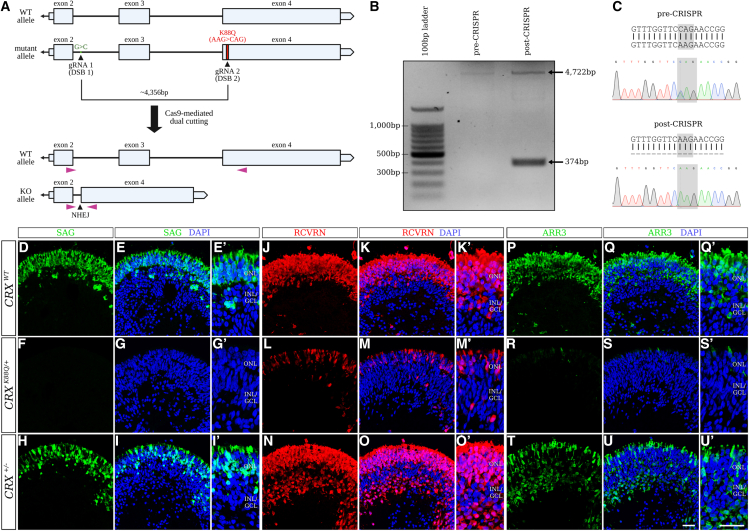
Cases of Leber congenital amaurosis caused by mutations in CRX (LCA7) exhibit an early form of the disease and show signs of significant photoreceptor dysfunction and eventual loss. To establish a translational in vitro model system to study gene-editing-based therapies, we generated LCA7 retinal organoids harboring a dominant disease-causing mutation in CRX. Our LCA7 retinal organoids develop signs of immature and dysfunctional photoreceptor cells, providing us with a reliable in vitro model to recapitulate LCA7. Furthermore, we performed a proof-of-concept study in which we utilize allele-specific CRISPR/Cas9-based gene editing to knock out mutant CRX and saw moderate rescue of photoreceptor phenotypes in our organoids. This work provides early evidence for an effective approach to treat LCA7, which can be applied more broadly to other dominant genetic diseases.
Keywords: CRISPR/Cas9; CRX; LCA7; Leber congenital amaurosis; allelic knockdown; gene editing; photoreceptors cells; retinal organoid; scRNA-seq.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Capowski E.E., Samimi K., Mayerl S.J., Phillips M.J., Pinilla I., Howden S.E., Saha J., Jansen A.D., Edwards K.L., Jager L.D., et al. Reproducibility and staging of 3D human retinal organoids across multiple pluripotent stem cell lines. Development. 2019;146 doi: 10.1242/dev.171686. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
