

Adaptation to an amoeba host drives selection of virulence-associated traits in Vibrio cholerae
- PMID: 34654895
- PMCID: PMC8857207
- DOI: 10.1038/s41396-021-01134-2
Adaptation to an amoeba host drives selection of virulence-associated traits in Vibrio cholerae
Abstract
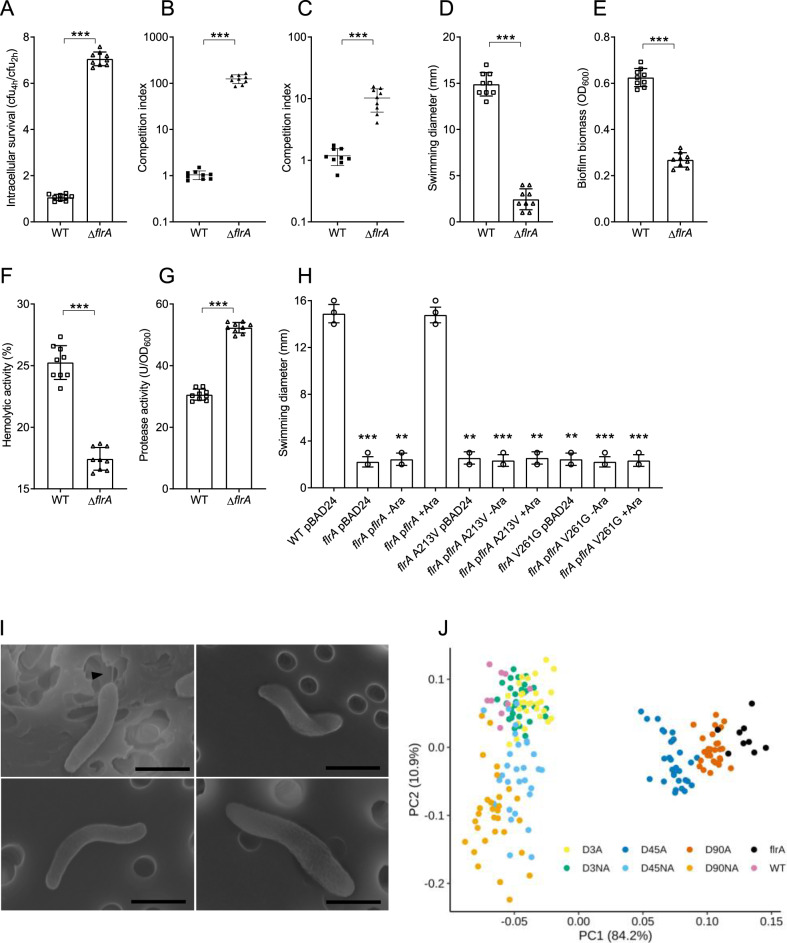
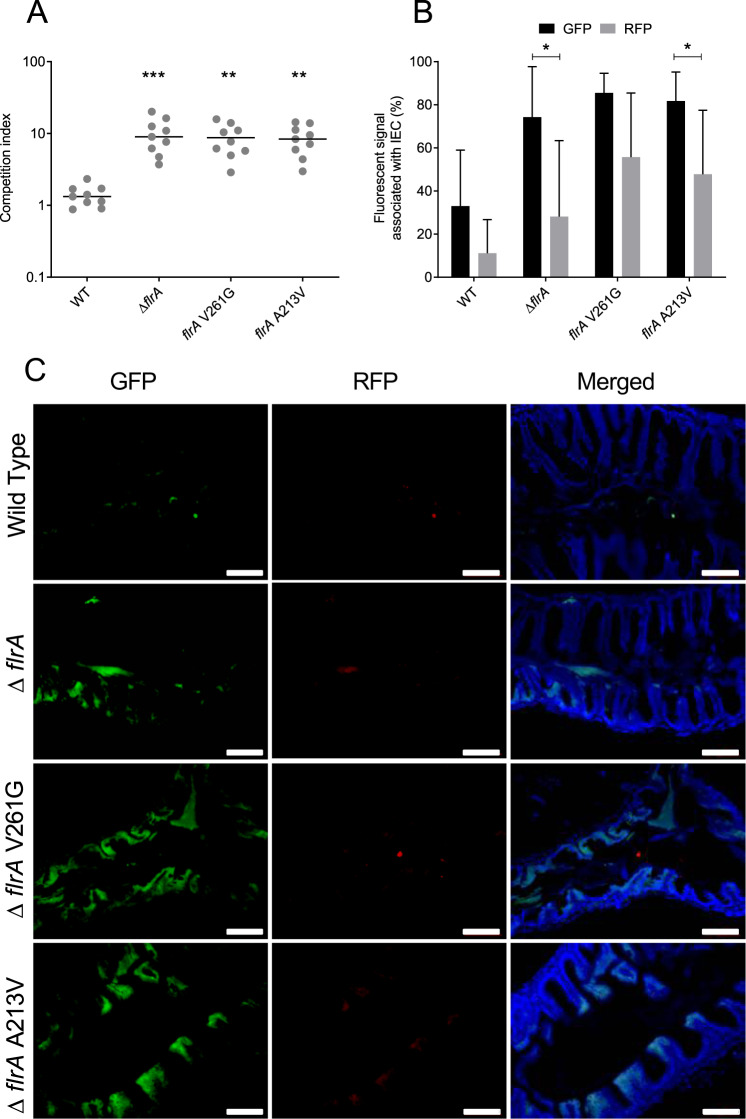
Predation by heterotrophic protists drives the emergence of adaptive traits in bacteria, and often these traits lead to altered interactions with hosts and persistence in the environment. Here we studied adaptation of the cholera pathogen, Vibrio cholerae during long-term co-incubation with the protist host, Acanthamoeba castellanii. We determined phenotypic and genotypic changes associated with long-term intra-amoebal host adaptation and how this impacts pathogen survival and fitness. We showed that adaptation to the amoeba host leads to temporal changes in multiple phenotypic traits in V. cholerae that facilitate increased survival and competitive fitness in amoeba. Genome sequencing and mutational analysis revealed that these altered lifestyles were linked to non-synonymous mutations in conserved regions of the flagellar transcriptional regulator, flrA. Additionally, the mutations resulted in enhanced colonisation in zebrafish, establishing a link between adaptation of V. cholerae to amoeba predation and enhanced environmental persistence. Our results show that pressure imposed by amoeba on V. cholerae selects for flrA mutations that serves as a key driver for adaptation. Importantly, this study provides evidence that adaptive traits that evolve in pathogens in response to environmental predatory pressure impact the colonisation of eukaryotic organisms by these pathogens.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures



References
-
- Jürgens K, Matz C. Predation as a shaping force for the phenotypic and genotypic composition of planktonic bacteria. Anton Van Leeuwen. 2002;81:413–34. - PubMed
-
- Brüssow H. Bacteria between protists and phages: from antipredation strategies to the evolution of pathogenicity. Mol Microbiol. 2007;65:583–9. - PubMed
-
- Pernthaler J. Predation on prokaryotes in the water column and its ecological implications. Nat Rev Microbiol. 2005;3:537–46. - PubMed
-
- Matz C, Kjelleberg S. Off the hook–how bacteria survive protozoan grazing. Trends Microbiol. 2005;13:302–7. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
