

OsRELA Regulates Leaf Inclination by Repressing the Transcriptional Activity of OsLIC in Rice
- PMID: 34659326
- PMCID: PMC8519309
- DOI: 10.3389/fpls.2021.760041
OsRELA Regulates Leaf Inclination by Repressing the Transcriptional Activity of OsLIC in Rice
Abstract
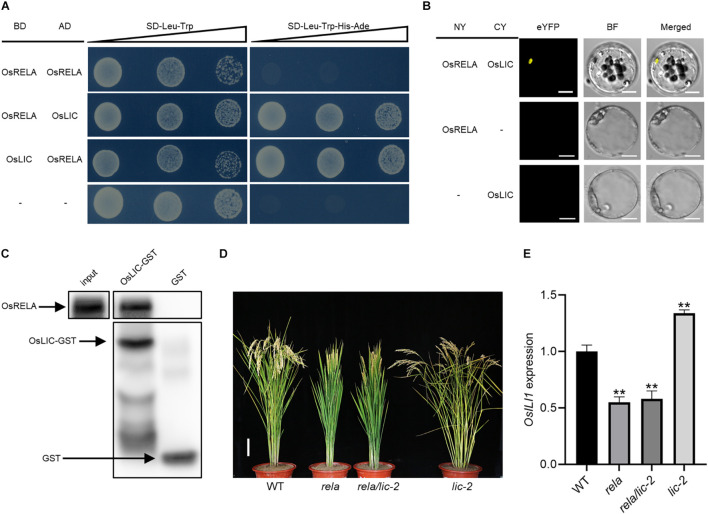
Leaf angle is one of the most important agronomic traits in rice, and changes in leaf angle can alter plant architecture to affect photosynthetic efficiency and thus determine grain yield. Therefore, it is important to identify key genes controlling leaf angle and elucidate the molecular mechanisms to improve rice yield. We obtained a mutant rela (regulator of leaf angle) with reduced leaf angle in rice by EMS mutagenesis, and map-based cloning revealed that OsRELA encodes a protein of unknown function. Coincidentally, DENSE AND ERECT PANICLE 2 (DEP2) was reported in a previous study with the same gene locus. RNA-seq analysis revealed that OsRELA is involved in regulating the expression of ILI and Expansin family genes. Biochemical and genetic analyses revealed that OsRELA is able to interact with OsLIC, a negative regulator of BR signaling, through its conserved C-terminal domain, which is essential for OsRELA function in rice. The binding of OsRELA can activate the expression of downstream genes repressed by OsLIC, such as OsILI1, a positive regulator of leaf inclination in rice. Therefore, our results suggest that OsRELA can act as a transcriptional regulator and is involved in the regulation of leaf inclination by regulating the transcriptional activity of OsLIC.
Keywords: OsLIC; OsRELA; leaf inclination; phytohormone; rice.
Copyright © 2021 Zhu, Xing, Teng, Deng, Shen, Ai, Lu, Zhang and Zhang.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures






References
-
- Bian H., Xie Y., Guo F., Han N., Ma S., Zeng Z., et al. (2012). Distinctive expression patterns and roles of the miRNA393/TIR1 homolog module in regulating flag leaf inclination and primary and crown root growth in rice (Oryza sativa). New Phytol. 196 149–161. 10.1111/j.1469-8137.2012.04248.x - DOI - PubMed
LinkOut - more resources
Full Text Sources