

Microglia control glutamatergic synapses in the adult mouse hippocampus
- PMID: 34661306
- PMCID: PMC9297980
- DOI: 10.1002/glia.24101
Microglia control glutamatergic synapses in the adult mouse hippocampus
Abstract
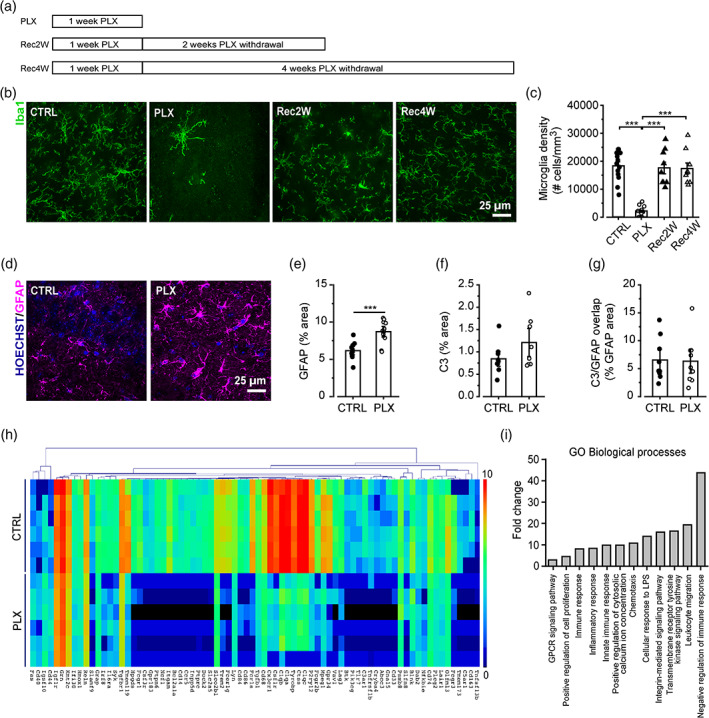
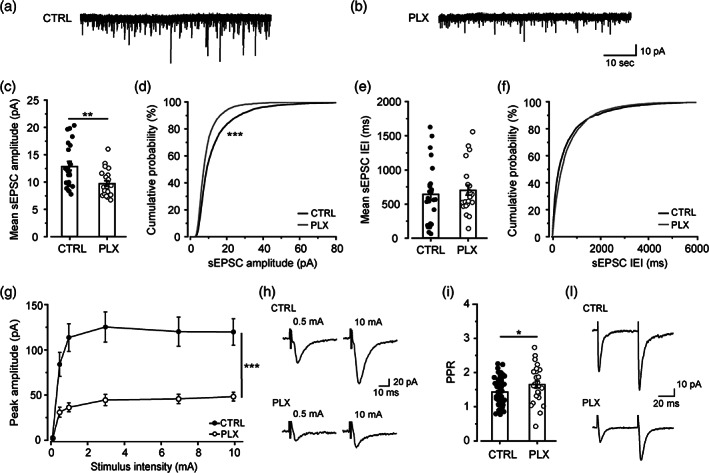
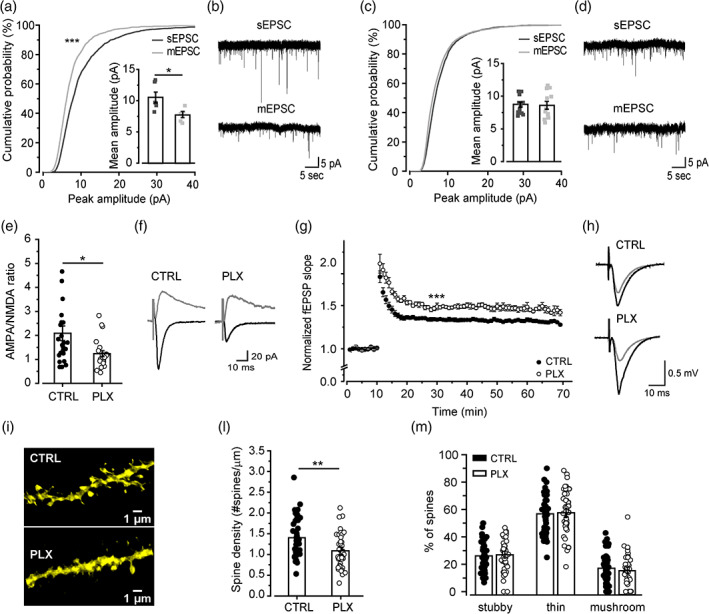
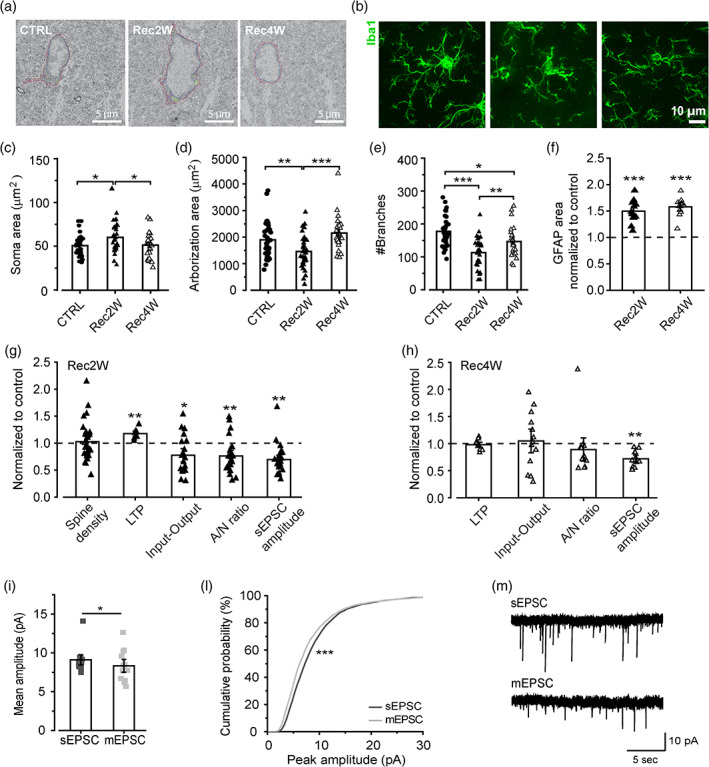
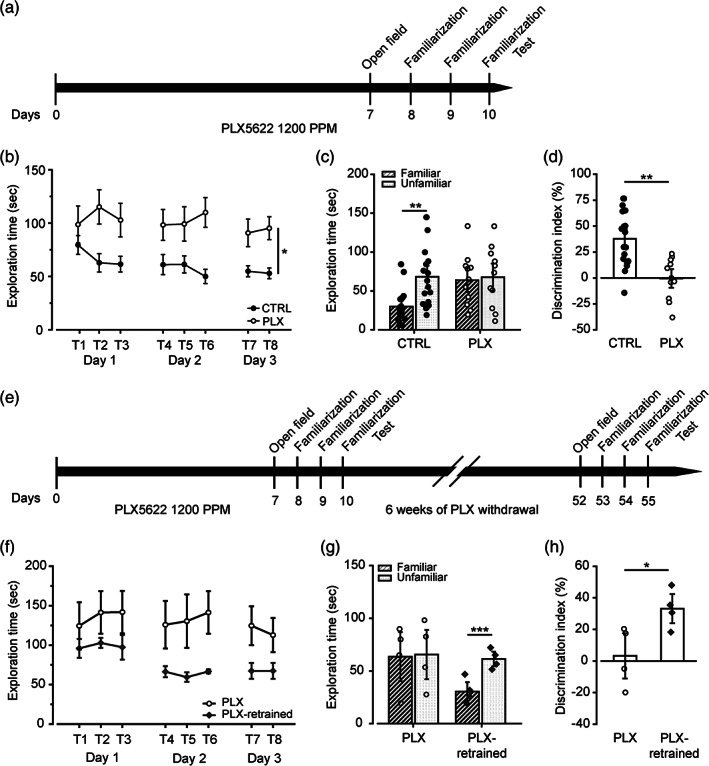
Microglia cells are active players in regulating synaptic development and plasticity in the brain. However, how they influence the normal functioning of synapses is largely unknown. In this study, we characterized the effects of pharmacological microglia depletion, achieved by administration of PLX5622, on hippocampal CA3-CA1 synapses of adult wild type mice. Following microglial depletion, we observed a reduction of spontaneous and evoked glutamatergic activity associated with a decrease of dendritic spine density. We also observed the appearance of immature synaptic features and higher levels of plasticity. Microglia depleted mice showed a deficit in the acquisition of the Novel Object Recognition task. These events were accompanied by hippocampal astrogliosis, although in the absence ofneuroinflammatory condition. PLX-induced synaptic changes were absent in Cx3cr1-/- mice, highlighting the role of CX3CL1/CX3CR1 axis in microglia control of synaptic functioning. Remarkably, microglia repopulation after PLX5622 withdrawal was associated with the recovery of hippocampal synapses and learning functions. Altogether, these data demonstrate that microglia contribute to normal synaptic functioning in the adult brain and that their removal induces reversible changes in organization and activity of glutamatergic synapses.
Keywords: glutamatergic transmission; hippocampus; learning; microglia; neuron-microglia interaction; synapses.
© 2021 The Authors. GLIA published by Wiley Periodicals LLC.
Conflict of interest statement
The authors declare that they have no conflict of interests.
Figures
References
-
- Akiyoshi, R. , Wake, H. , Kato, D. , Horiuchi, H. , Ono, R. , Ikegami, A. , Haruwaka, K. , Omori, T. , Tachibana, Y. , Moorhouse, A. J. , & Nabekura, J. (2018). Microglia Enhance Synapse Activity to Promote Local Network Synchronization. eNeuro, 5(5), 88–106. 10.1523/ENEURO.0088-18.2018 - DOI - PMC - PubMed
-
- Badimon, A. , Strasburger, H. J. , Ayata, P. , Chen, X. , Nair, A. , Ikegami, A. , Hwang, P. , Chan, A. T. , Graves, S. M. , Uweru, J. O. , Ledderose, C. , Kutlu, M. G. , Wheeler, M. A. , Kahan, A. , Ishikawa, M. , Wang, Y. C. , Loh, Y. H. E. , Jiang, J. X. , Surmeier, D. J. , … Schaefer, A. (2020). Negative feedback control of neuronal activity by microglia. Nature, 10, 1–7. 10.1038/s41586-020-2777-8 - DOI - PMC - PubMed
-
- Barnes, C. A. , Jung, M. W. , McNaughton, B. L. , Korol, D. L. , Andreasson, K. , & Worley, P. F. (1994). LTP saturation and spatial learning disruption: Effects of task variables and saturation levels. Journal of Neuroscience, 14(10), 5793–5806. 10.1523/jneurosci.14-10-05793.1994 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
