

Comprehensive multi-omics integration identifies differentially active enhancers during human brain development with clinical relevance
- PMID: 34663447
- PMCID: PMC8524963
- DOI: 10.1186/s13073-021-00980-1
Comprehensive multi-omics integration identifies differentially active enhancers during human brain development with clinical relevance
Abstract
Background: Non-coding regulatory elements (NCREs), such as enhancers, play a crucial role in gene regulation, and genetic aberrations in NCREs can lead to human disease, including brain disorders. The human brain is a complex organ that is susceptible to numerous disorders; many of these are caused by genetic changes, but a multitude remain currently unexplained. Understanding NCREs acting during brain development has the potential to shed light on previously unrecognized genetic causes of human brain disease. Despite immense community-wide efforts to understand the role of the non-coding genome and NCREs, annotating functional NCREs remains challenging.
Methods: Here we performed an integrative computational analysis of virtually all currently available epigenome data sets related to human fetal brain.
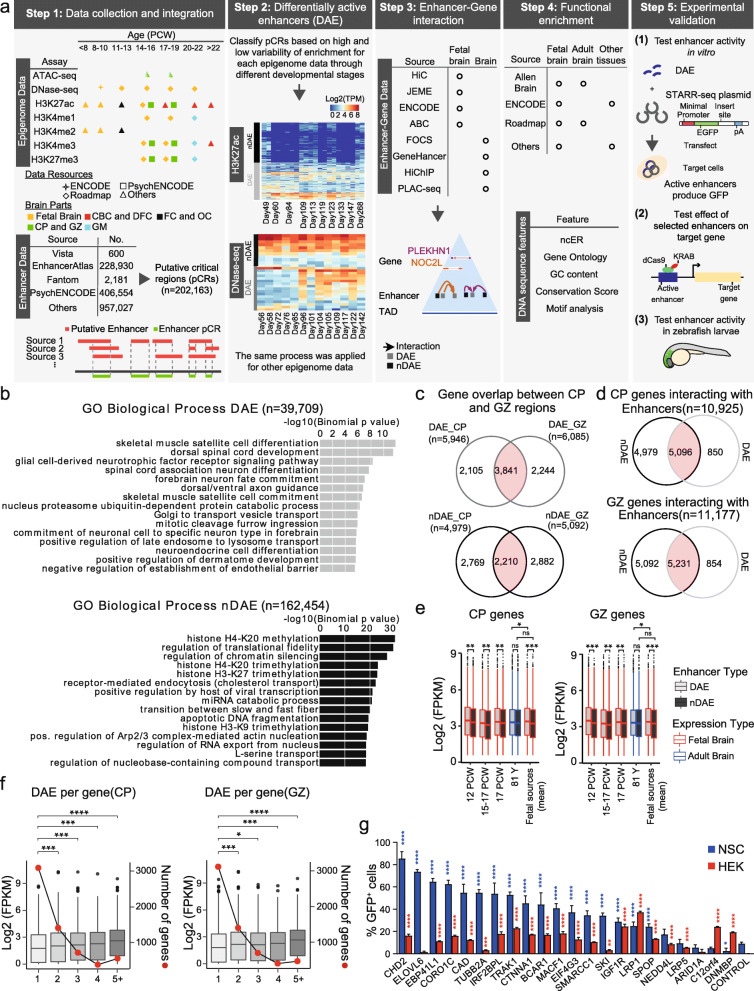
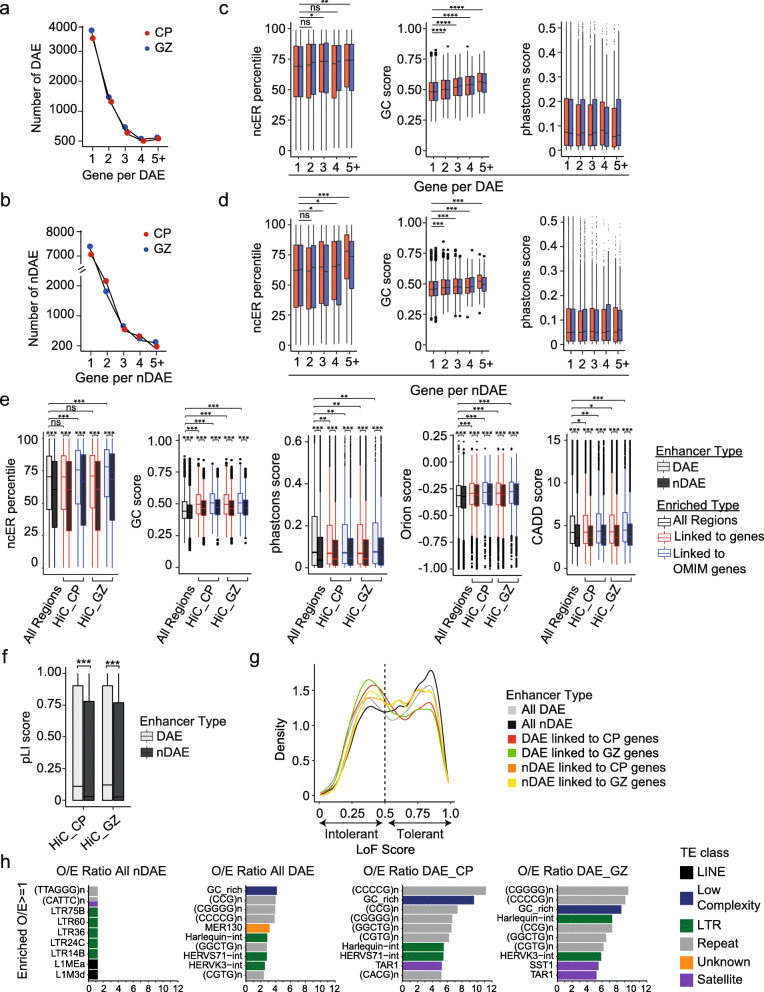
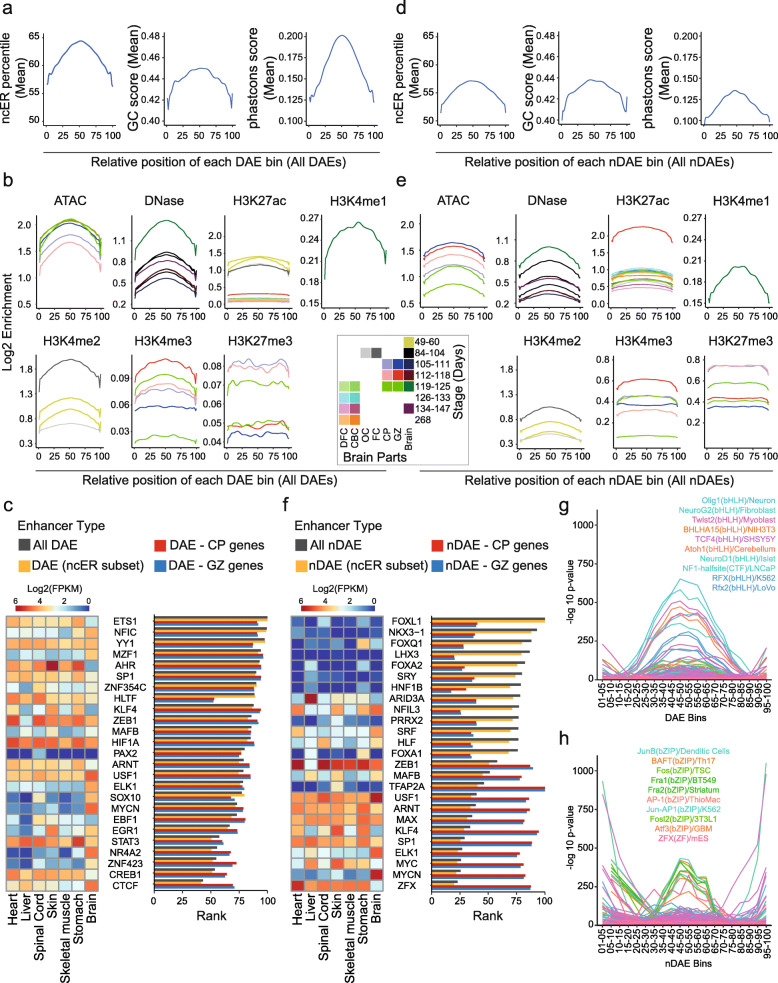
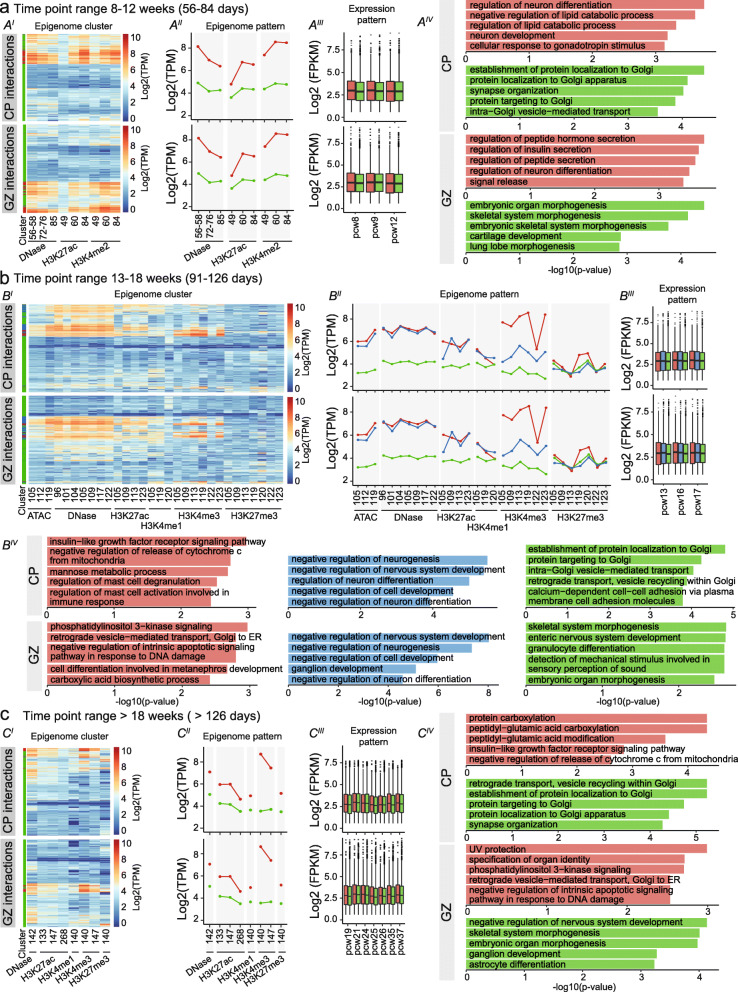
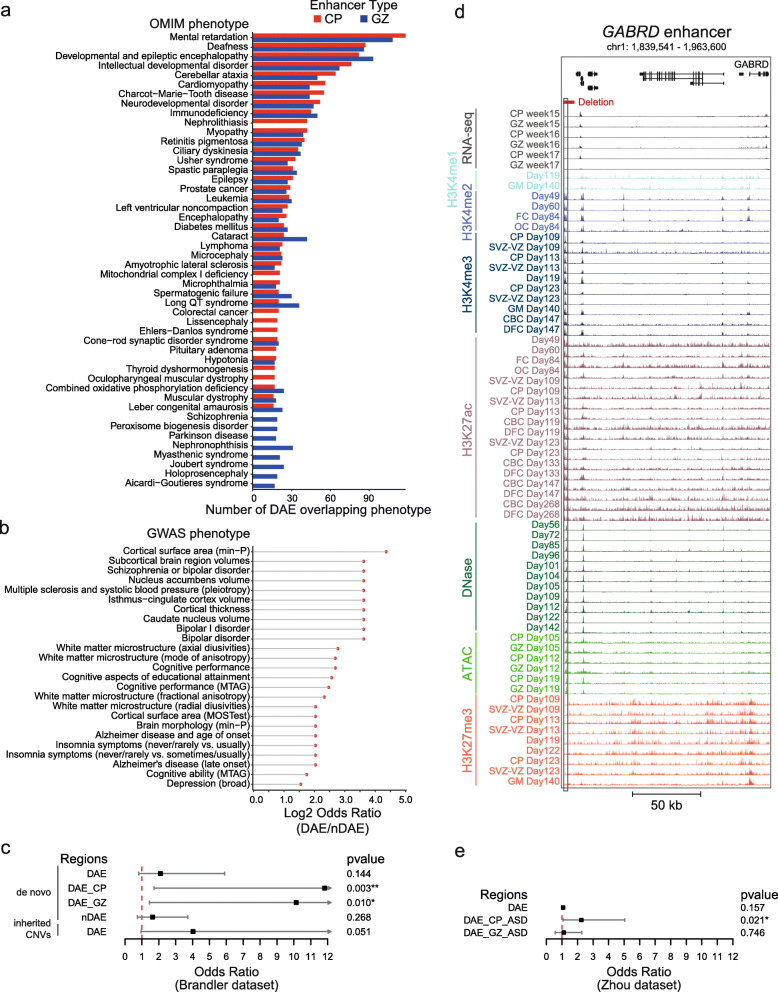
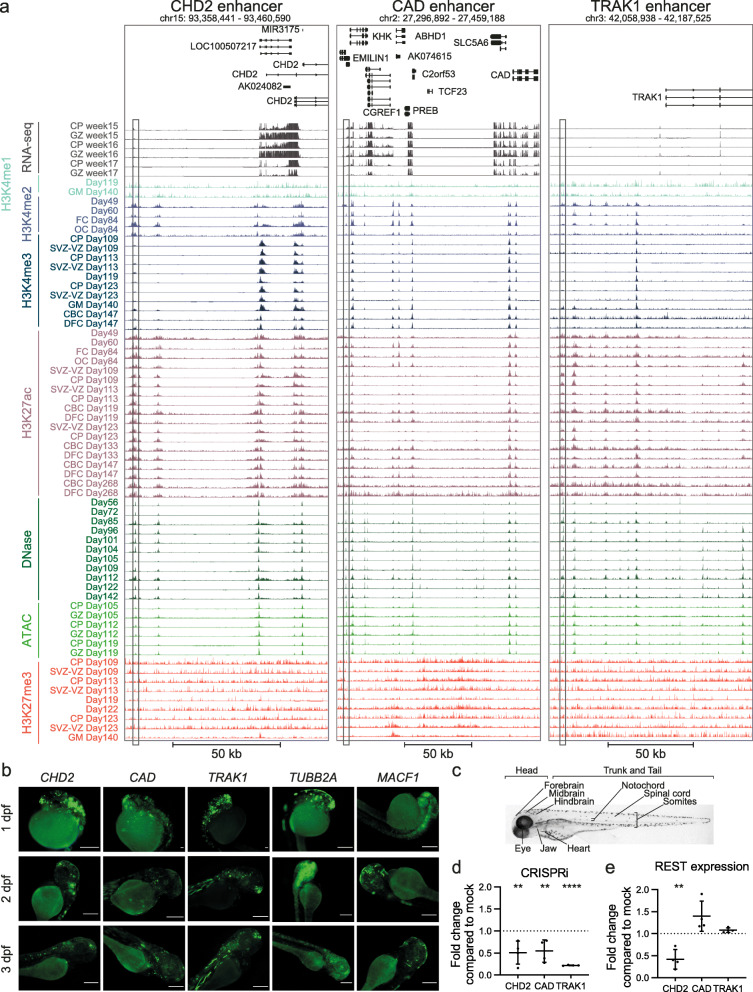
Results: Our in-depth analysis unravels 39,709 differentially active enhancers (DAEs) that show dynamic epigenomic rearrangement during early stages of human brain development, indicating likely biological function. Many of these DAEs are linked to clinically relevant genes, and functional validation of selected DAEs in cell models and zebrafish confirms their role in gene regulation. Compared to enhancers without dynamic epigenomic rearrangement, DAEs are subjected to higher sequence constraints in humans, have distinct sequence characteristics and are bound by a distinct transcription factor landscape. DAEs are enriched for GWAS loci for brain-related traits and for genetic variation found in individuals with neurodevelopmental disorders, including autism.
Conclusion: This compendium of high-confidence enhancers will assist in deciphering the mechanism behind developmental genetics of human brain and will be relevant to uncover missing heritability in human genetic brain disorders.
Keywords: Clinical genetics; Computational analysis; Data integration; Enhancer; Epigenome; Gene regulatory elements; Human brain development; Mendelian disorders; Meta-analysis; Non-coding genome.
© 2021. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
- R01 MH094714/MH/NIMH NIH HHS/United States
- U01 MH103365/MH/NIMH NIH HHS/United States
- R01 MH100027/MH/NIMH NIH HHS/United States
- U01 MH103392/MH/NIMH NIH HHS/United States
- RC2 MH090047/MH/NIMH NIH HHS/United States
- U01 MH103346/MH/NIMH NIH HHS/United States
- R01 MH105472/MH/NIMH NIH HHS/United States
- U01 MH103340/MH/NIMH NIH HHS/United States
- U01 MH103339/MH/NIMH NIH HHS/United States
- R21 MH105881/MH/NIMH NIH HHS/United States
- U01 ES017166/ES/NIEHS NIH HHS/United States
- P50 MH106934/MH/NIMH NIH HHS/United States
- U01 ES017154/ES/NIEHS NIH HHS/United States
- R21 MH103877/MH/NIMH NIH HHS/United States
- U01 DA025956/DA/NIDA NIH HHS/United States
- R01 MH110927/MH/NIMH NIH HHS/United States
- U01 MH081896/MH/NIMH NIH HHS/United States
- R01 MH060233/MH/NIMH NIH HHS/United States
- U01 ES017156/ES/NIEHS NIH HHS/United States
- R21 MH102791/MH/NIMH NIH HHS/United States
- R01 MH105898/MH/NIMH NIH HHS/United States
- RC2 MH089929/MH/NIMH NIH HHS/United States
- U01 ES017155/ES/NIEHS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
