

Enhanced IL-2 in early life limits the development of TFH and protective antiviral immunity
- PMID: 34665220
- PMCID: PMC8529914
- DOI: 10.1084/jem.20201555
Enhanced IL-2 in early life limits the development of TFH and protective antiviral immunity
Abstract
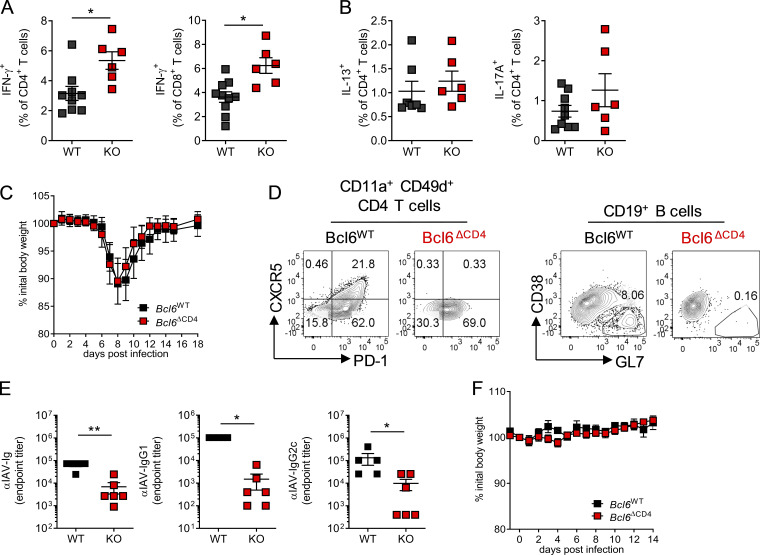
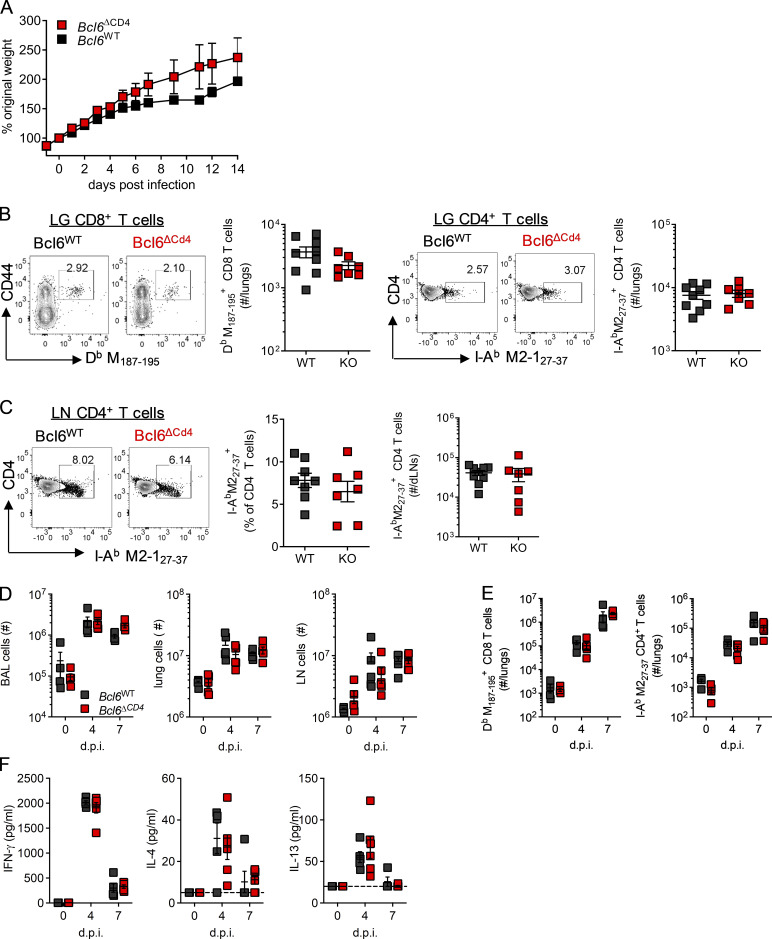
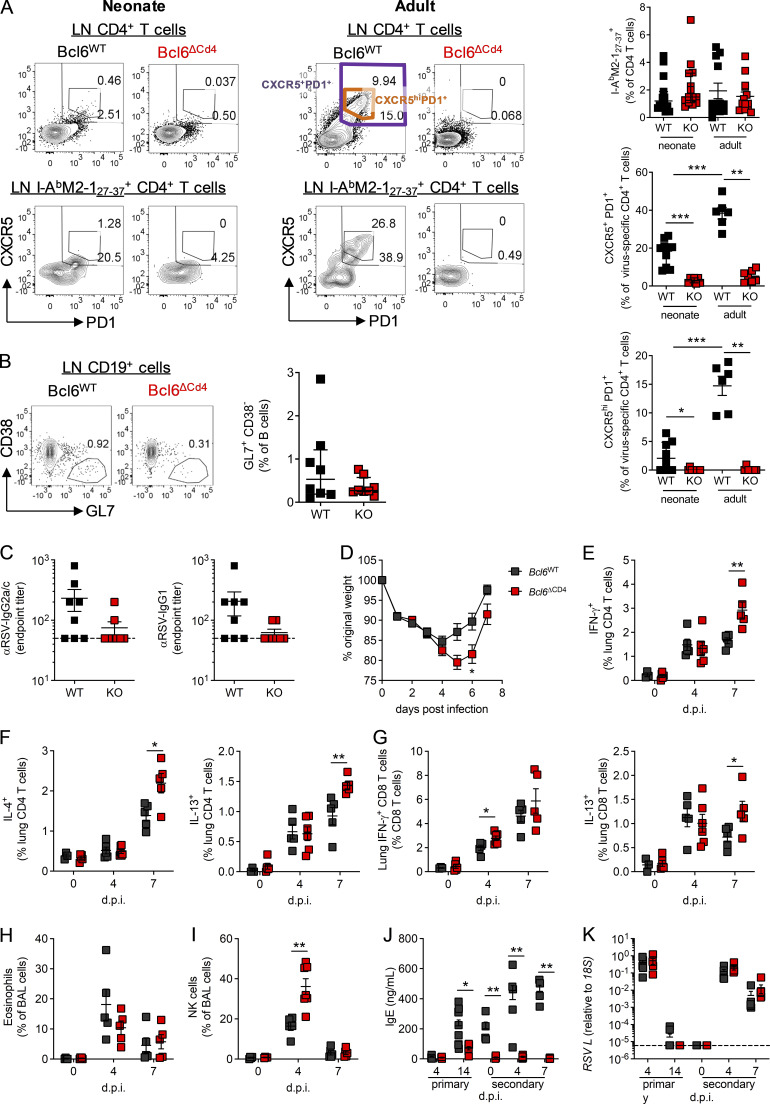
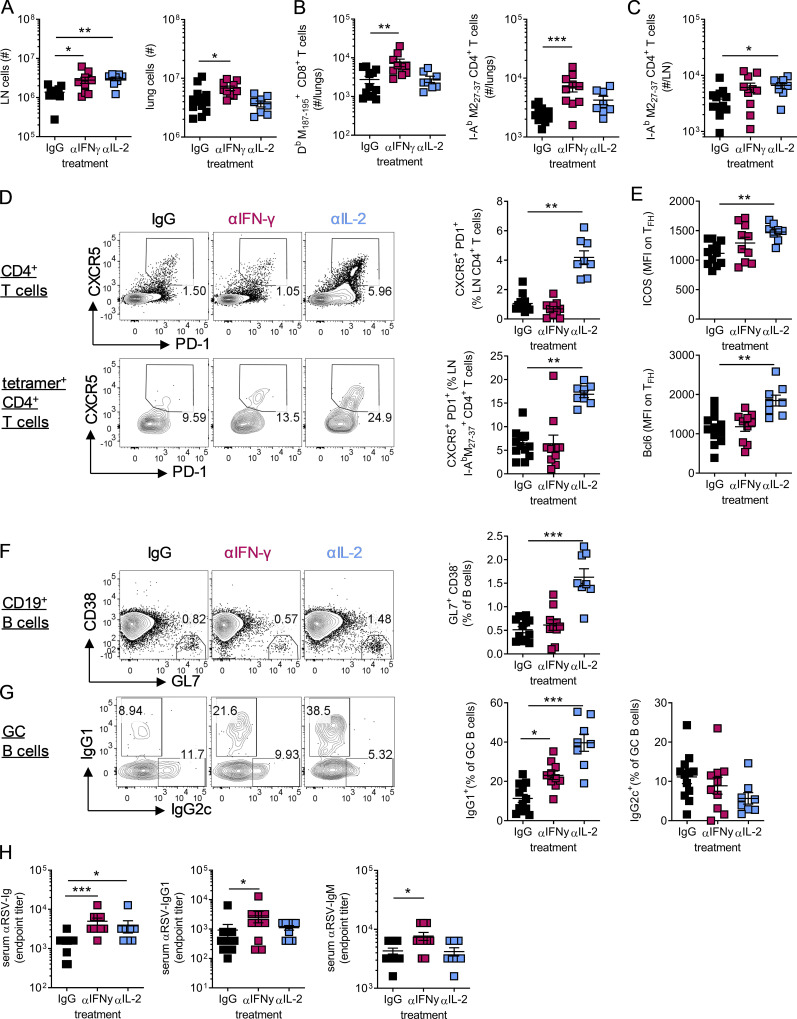
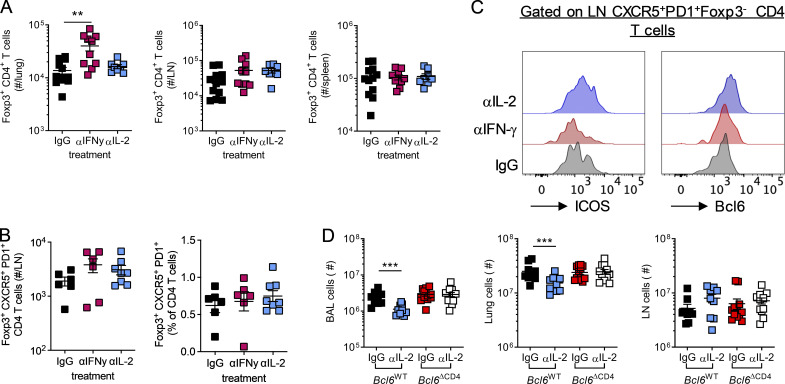
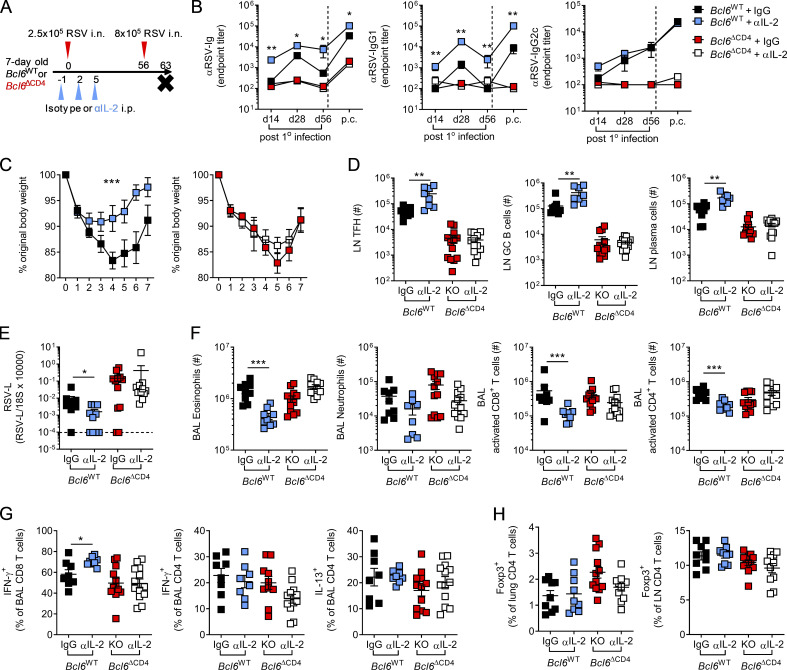
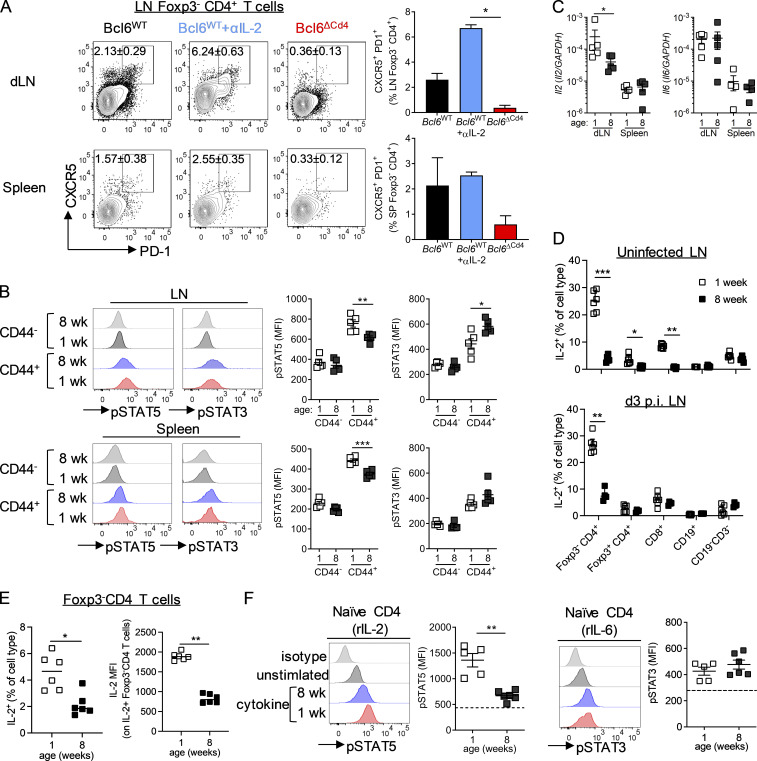
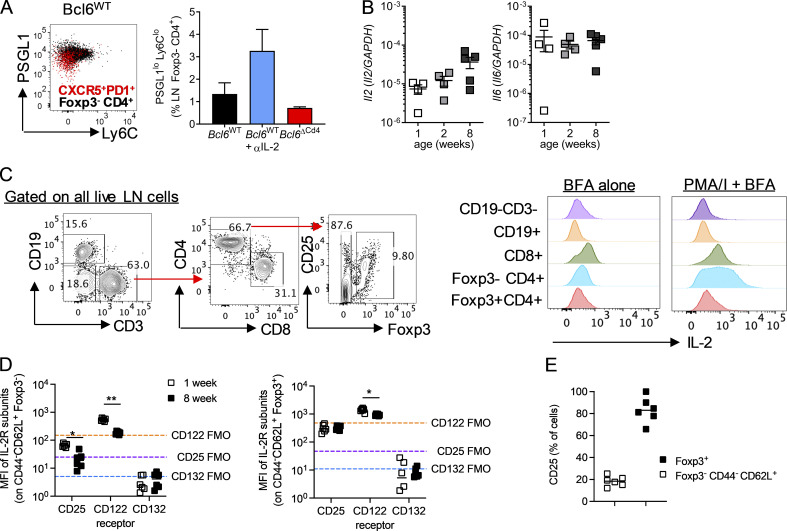
T follicular helper cell (TFH)-dependent antibody responses are critical for long-term immunity. Antibody responses are diminished in early life, limiting long-term protective immunity and allowing prolonged or recurrent infection, which may be important for viral lung infections that are highly prevalent in infancy. In a murine model using respiratory syncytial virus (RSV), we show that TFH and the high-affinity antibody production they promote are vital for preventing disease on RSV reinfection. Following a secondary RSV infection, TFH-deficient mice had significantly exacerbated disease characterized by delayed viral clearance, increased weight loss, and immunopathology. TFH generation in early life was compromised by heightened IL-2 and STAT5 signaling in differentiating naive T cells. Neutralization of IL-2 during early-life RSV infection resulted in a TFH-dependent increase in antibody-mediated immunity and was sufficient to limit disease severity upon reinfection. These data demonstrate the importance of TFH in protection against recurrent RSV infection and highlight a mechanism by which this is suppressed in early life.
© 2021 Pyle et al.
Conflict of interest statement
Disclosures: H.T. Groves reported, "I am currently employed by the Wellcome Trust but I was not employed by the Wellcome Trust at the time of generating data for this research paper. I did not and do not have any role in the funding decisions behind the research included in the submitted work." J.S. Tregoning reported grants from MRC during the conduct of the study. No other disclosures were reported.
Figures
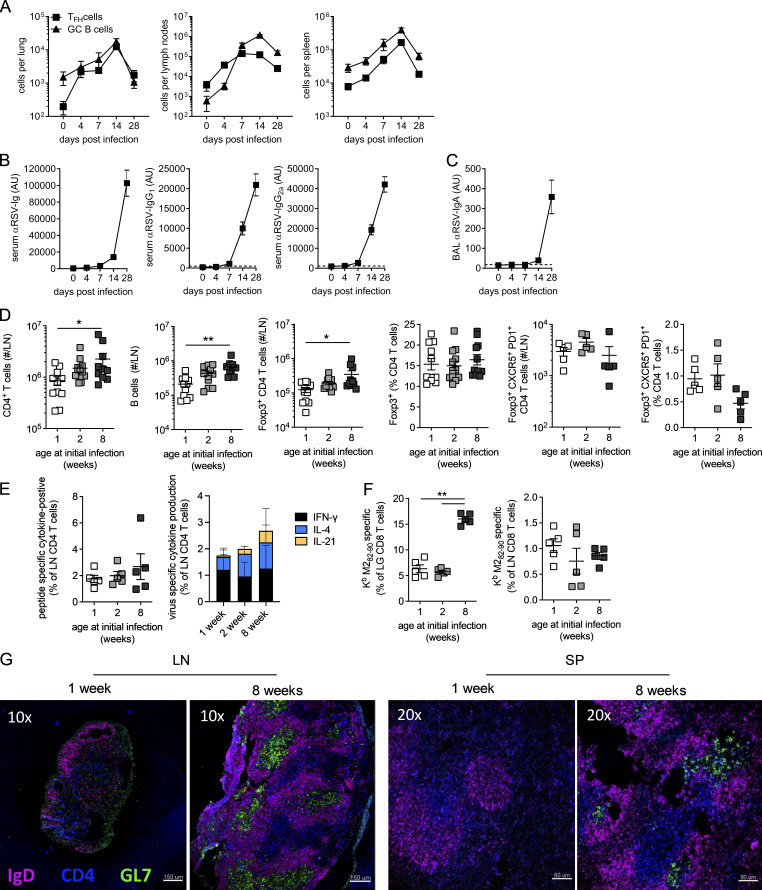
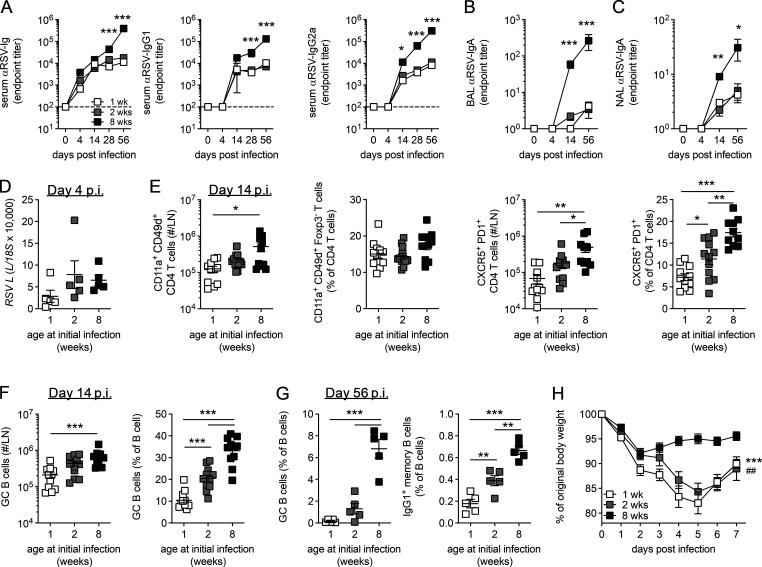
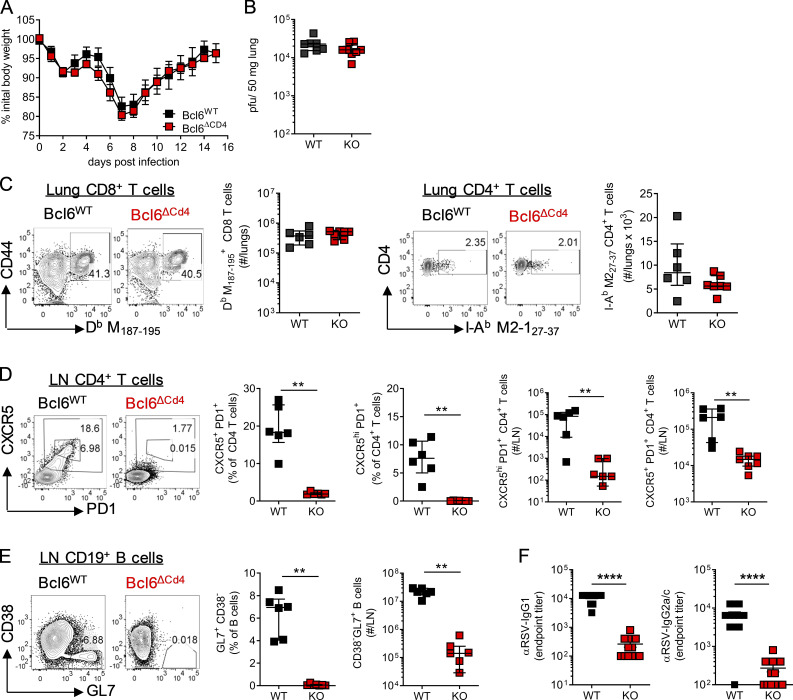
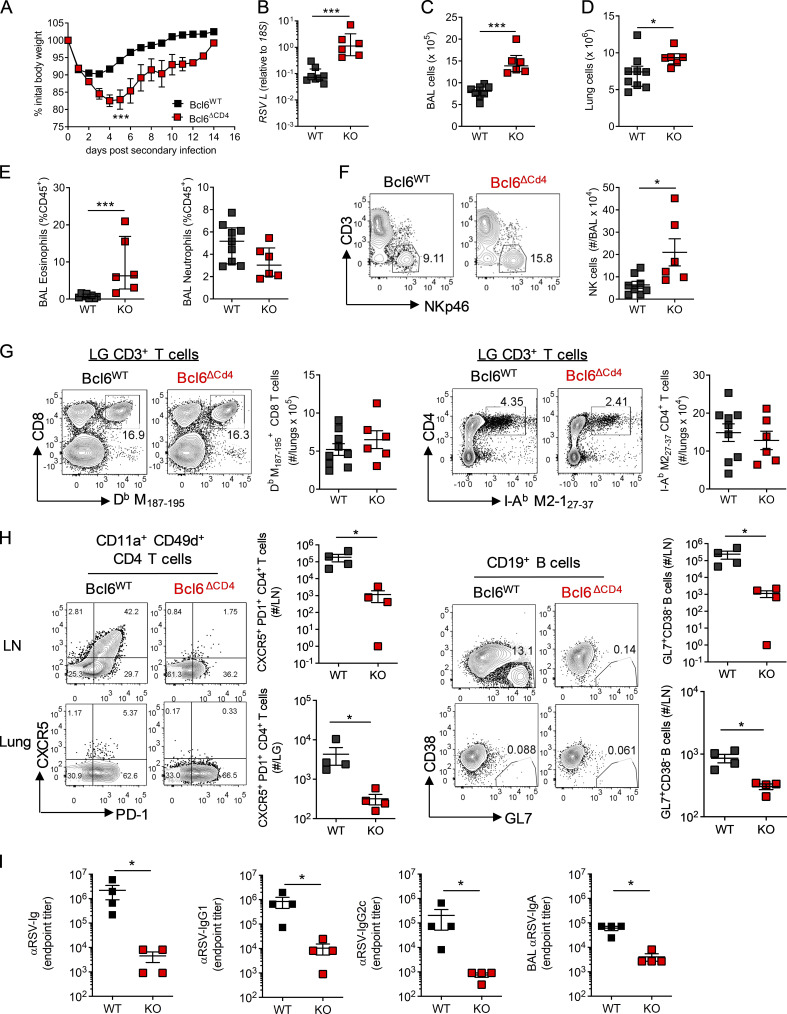
References
-
- Choi, Y.S., Kageyama R., Eto D., Escobar T.C., Johnston R.J., Monticelli L., Lao C., and Crotty S.. 2011. ICOS receptor instructs T follicular helper cell versus effector cell differentiation via induction of the transcriptional repressor Bcl6. Immunity. 34:932–946. 10.1016/j.immuni.2011.03.023 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
