

Regulation of intestinal immunity and tissue repair by enteric glia
- PMID: 34671159
- PMCID: PMC7612231
- DOI: 10.1038/s41586-021-04006-z
Regulation of intestinal immunity and tissue repair by enteric glia
Abstract
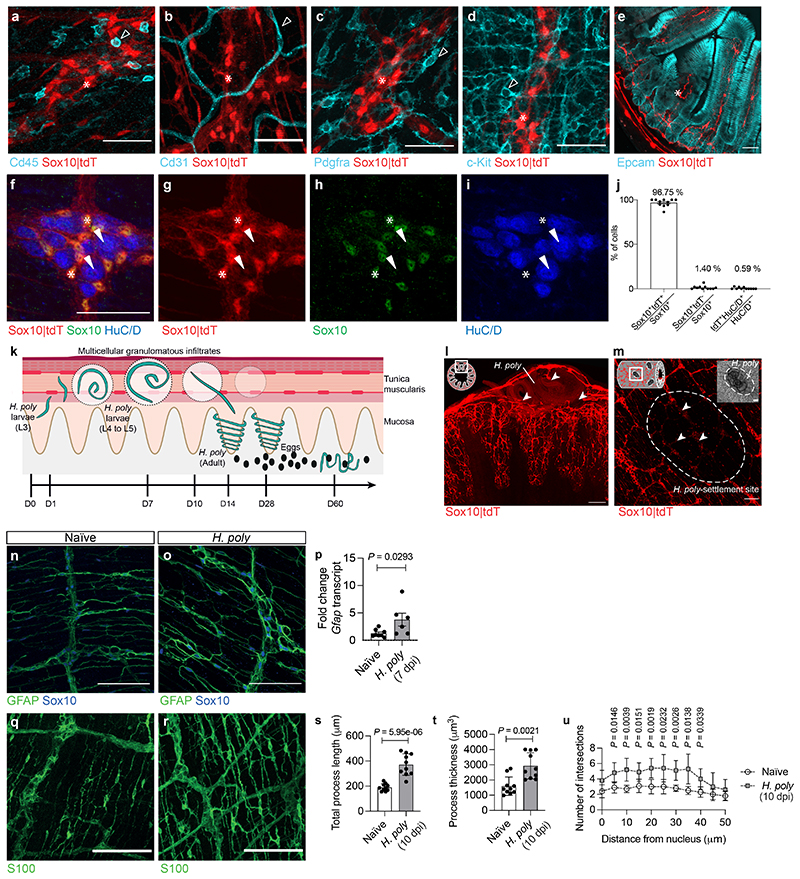
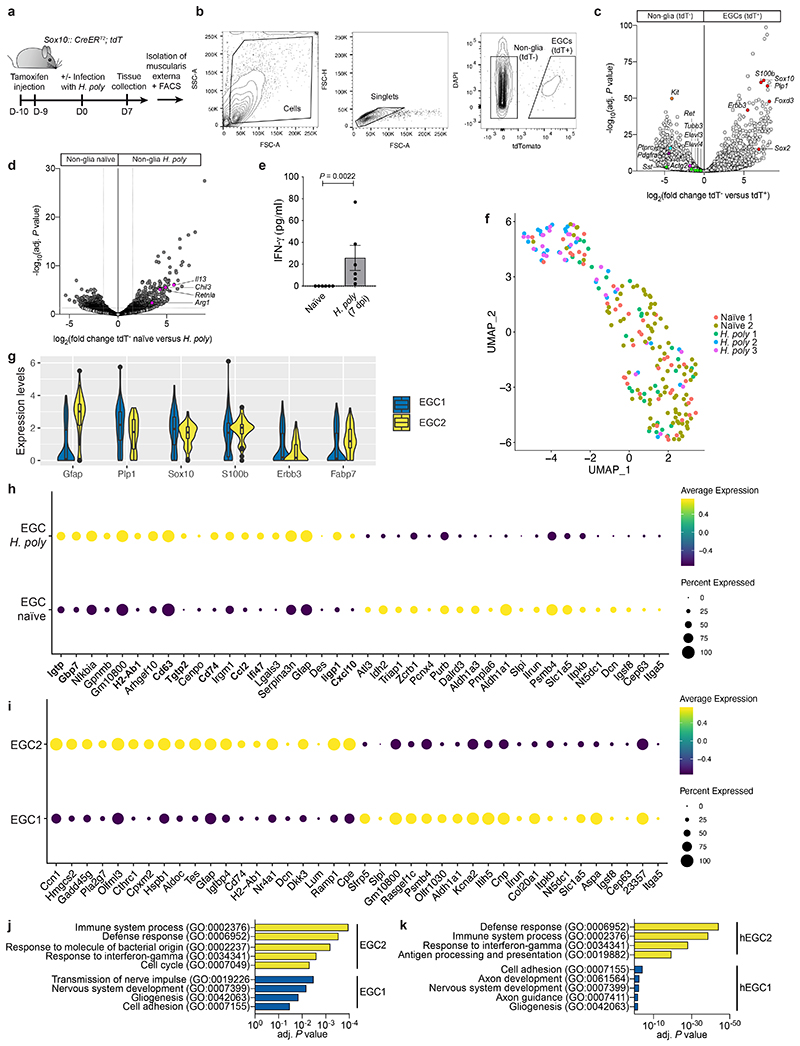
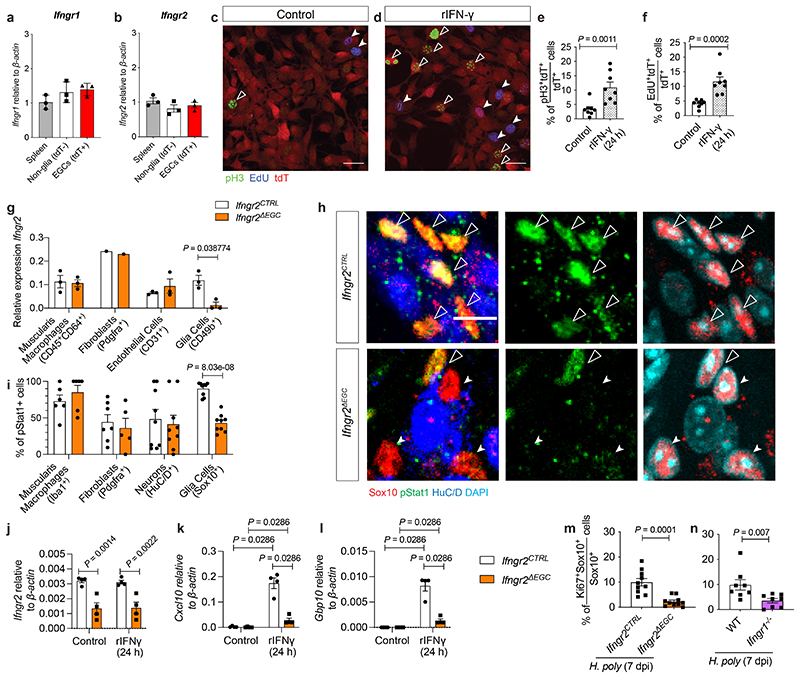
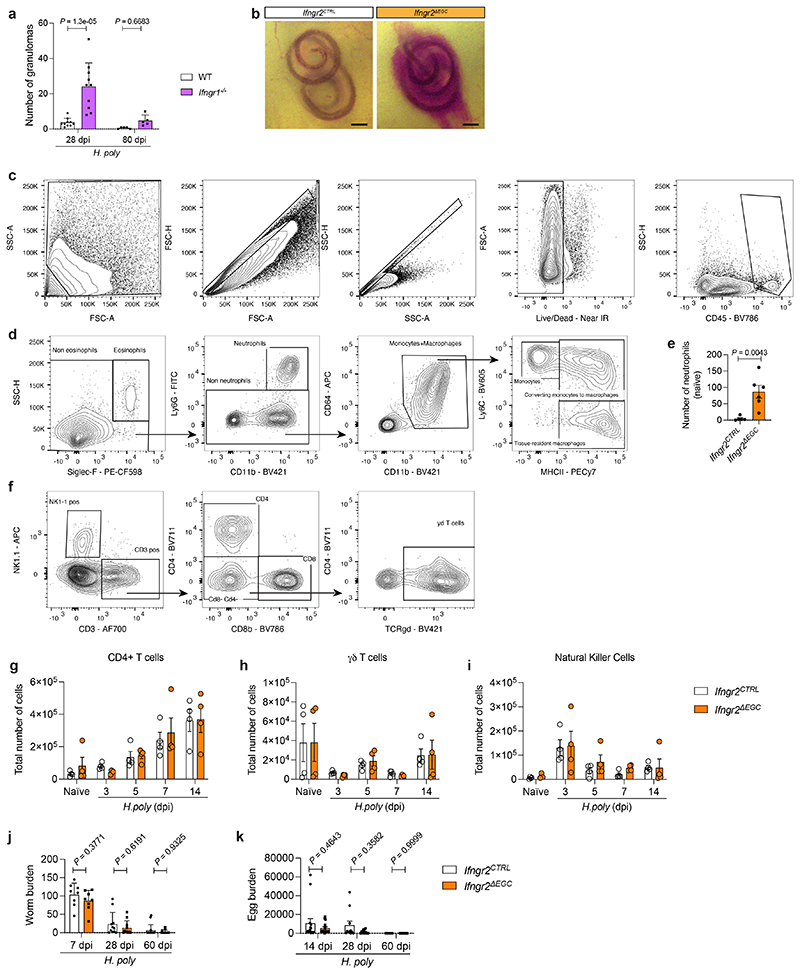
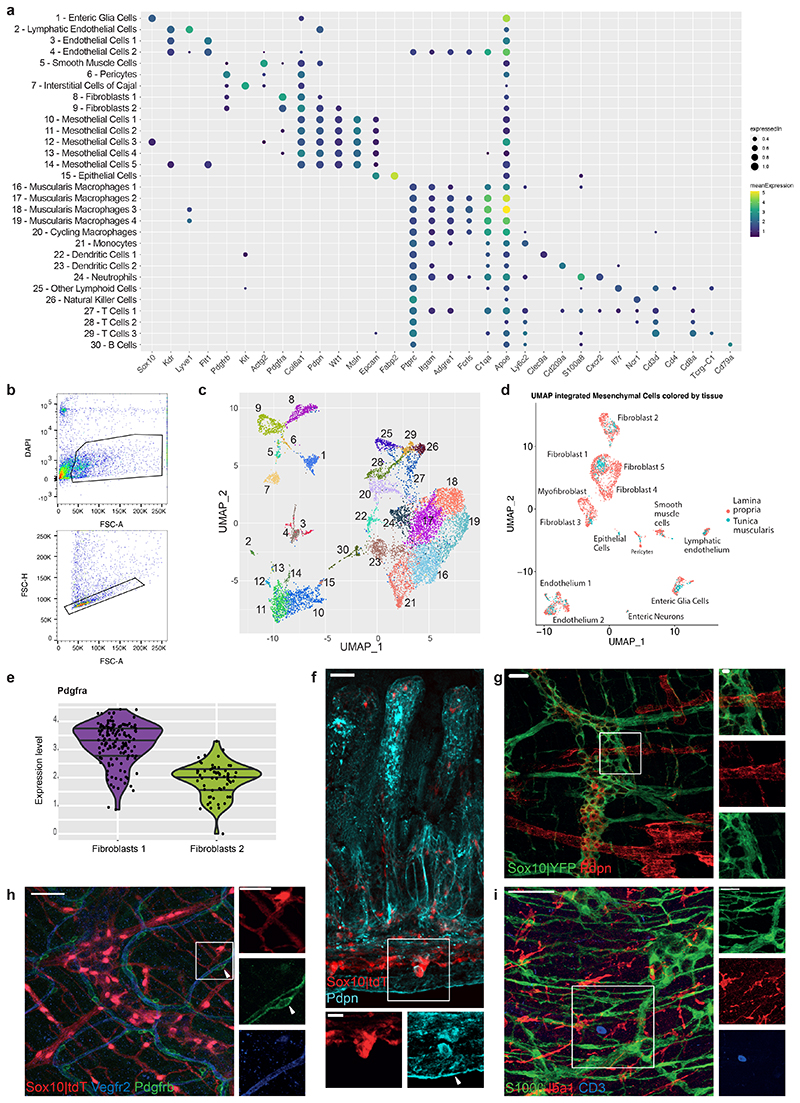
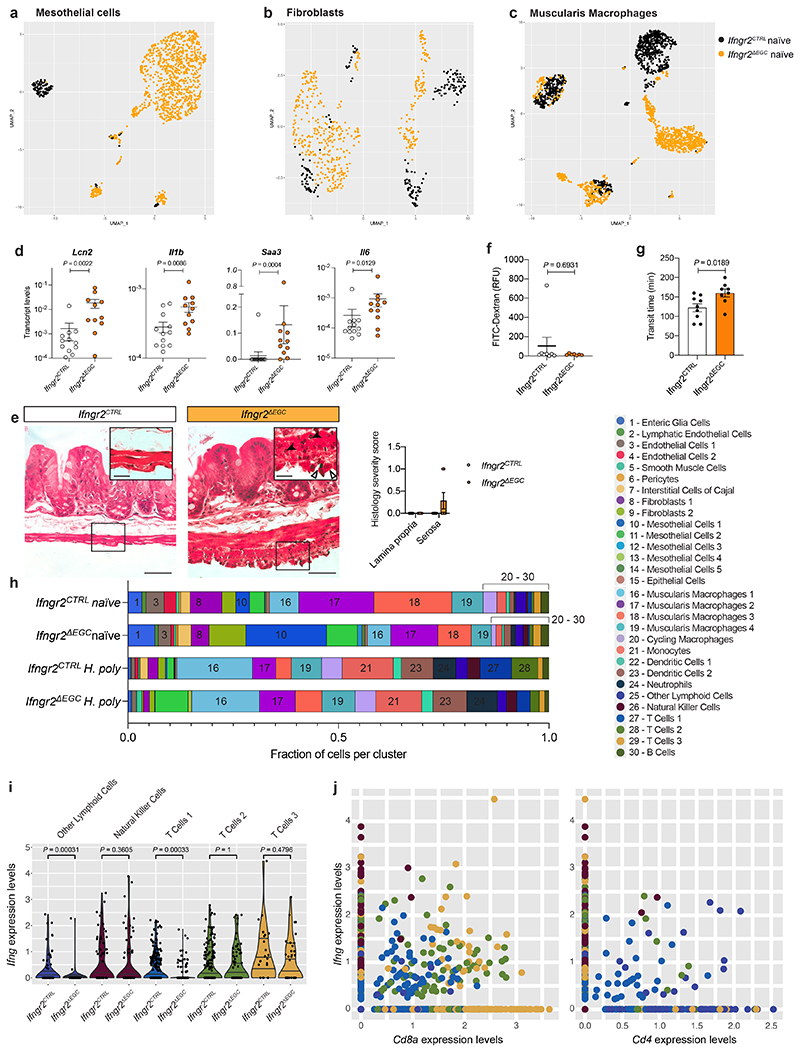
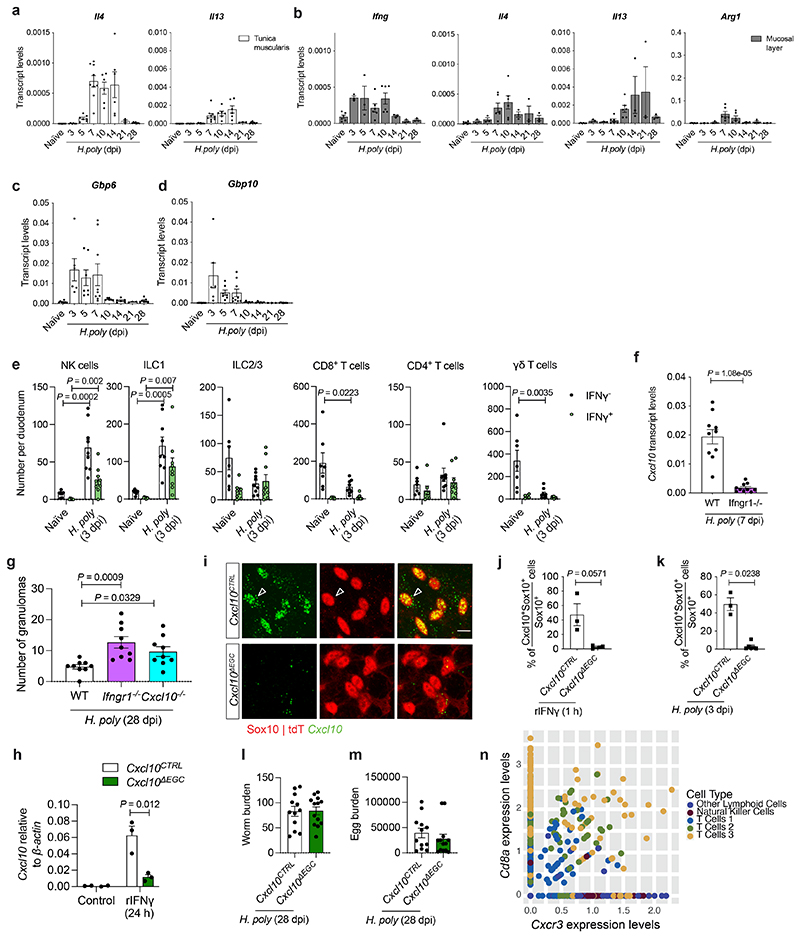
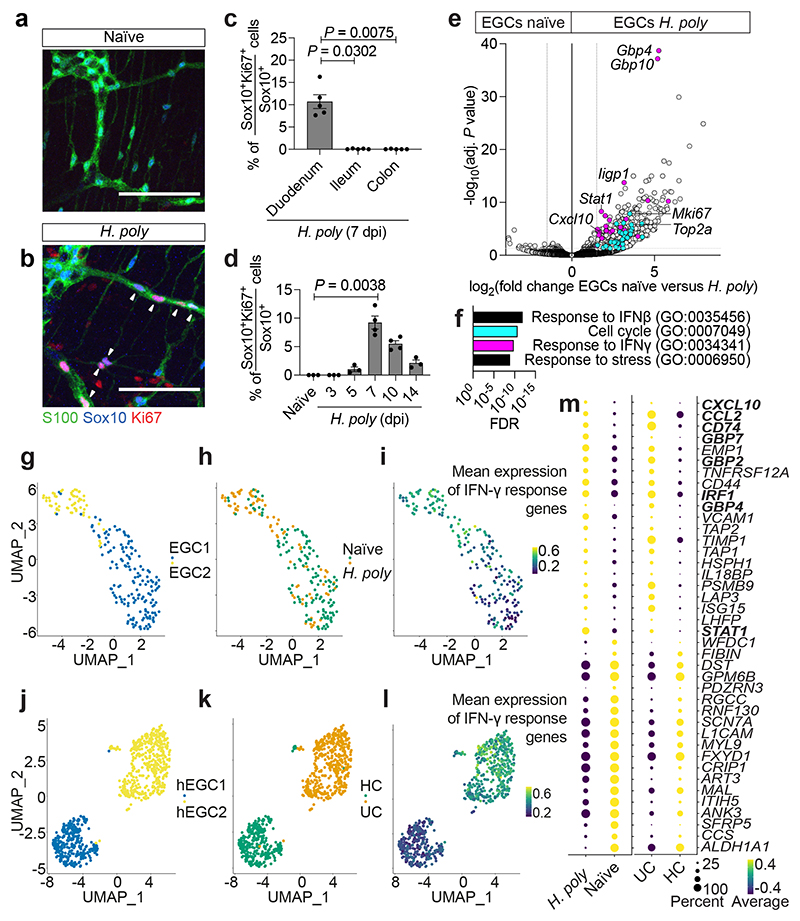
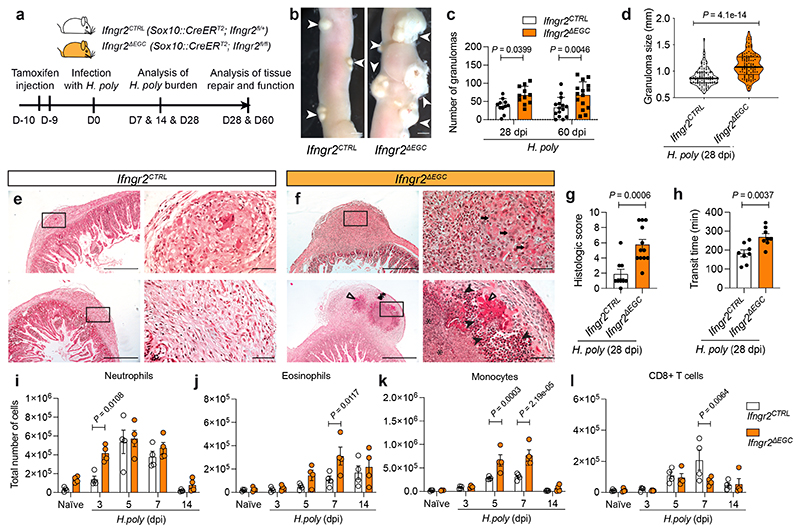
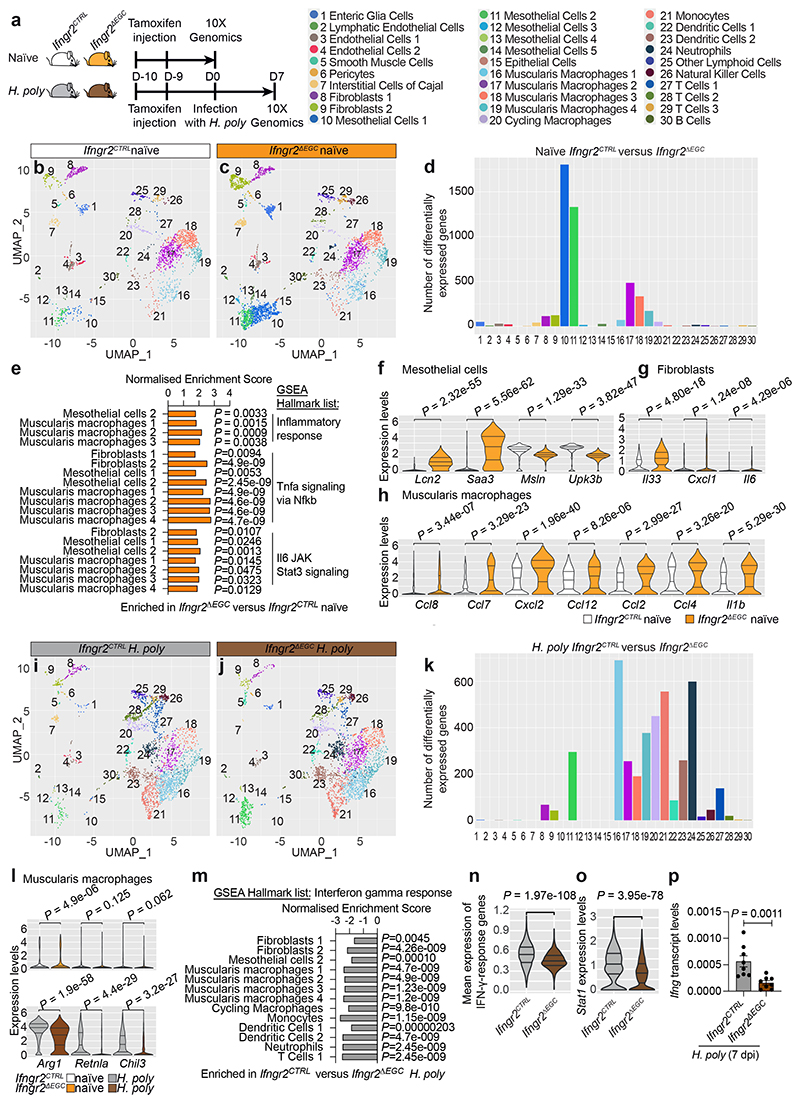
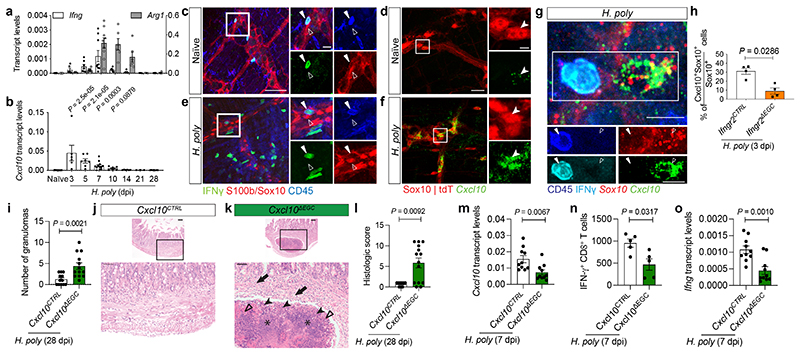
Tissue maintenance and repair depend on the integrated activity of multiple cell types1. Whereas the contributions of epithelial2,3, immune4,5 and stromal cells6,7 in intestinal tissue integrity are well understood, the role of intrinsic neuroglia networks remains largely unknown. Here we uncover important roles of enteric glial cells (EGCs) in intestinal homeostasis, immunity and tissue repair. We demonstrate that infection of mice with Heligmosomoides polygyrus leads to enteric gliosis and the upregulation of an interferon gamma (IFNγ) gene signature. IFNγ-dependent gene modules were also induced in EGCs from patients with inflammatory bowel disease8. Single-cell transcriptomics analysis of the tunica muscularis showed that glia-specific abrogation of IFNγ signalling leads to tissue-wide activation of pro-inflammatory transcriptional programs. Furthermore, disruption of the IFNγ-EGC signalling axis enhanced the inflammatory and granulomatous response of the tunica muscularis to helminths. Mechanistically, we show that the upregulation of Cxcl10 is an early immediate response of EGCs to IFNγ signalling and provide evidence that this chemokine and the downstream amplification of IFNγ signalling in the tunica muscularis are required for a measured inflammatory response to helminths and resolution of the granulomatous pathology. Our study demonstrates that IFNγ signalling in enteric glia is central to intestinal homeostasis and reveals critical roles of the IFNγ-EGC-CXCL10 axis in immune response and tissue repair after infectious challenge.
© 2021. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
Comment in
-
Glia mend the gut.Nat Rev Immunol. 2021 Dec;21(12):760-761. doi: 10.1038/s41577-021-00651-7. Nat Rev Immunol. 2021. PMID: 34725459 No abstract available.
-
Immunoregulatory role of enteric glia.Nat Rev Gastroenterol Hepatol. 2021 Dec;18(12):832-833. doi: 10.1038/s41575-021-00542-1. Nat Rev Gastroenterol Hepatol. 2021. PMID: 34725497 No abstract available.
-
Enteric glia worm their way into gut immunity.Immunity. 2021 Dec 14;54(12):2698-2700. doi: 10.1016/j.immuni.2021.11.014. Immunity. 2021. PMID: 34910939
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
