

Theta oscillations in rat infralimbic cortex are associated with the inhibition of cocaine seeking during extinction
- PMID: 34672059
- PMCID: PMC8922975
- DOI: 10.1111/adb.13106
Theta oscillations in rat infralimbic cortex are associated with the inhibition of cocaine seeking during extinction
Abstract
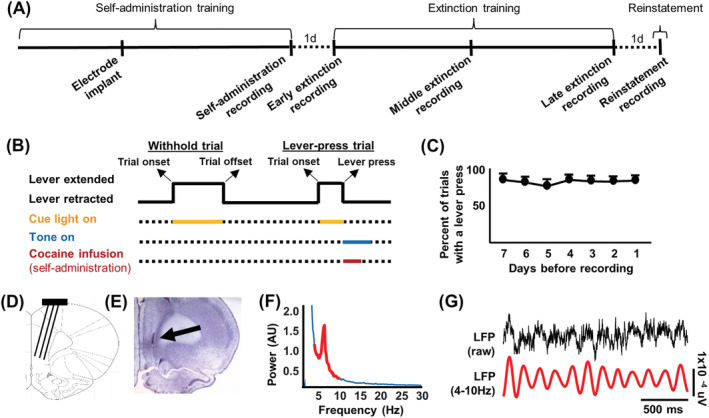
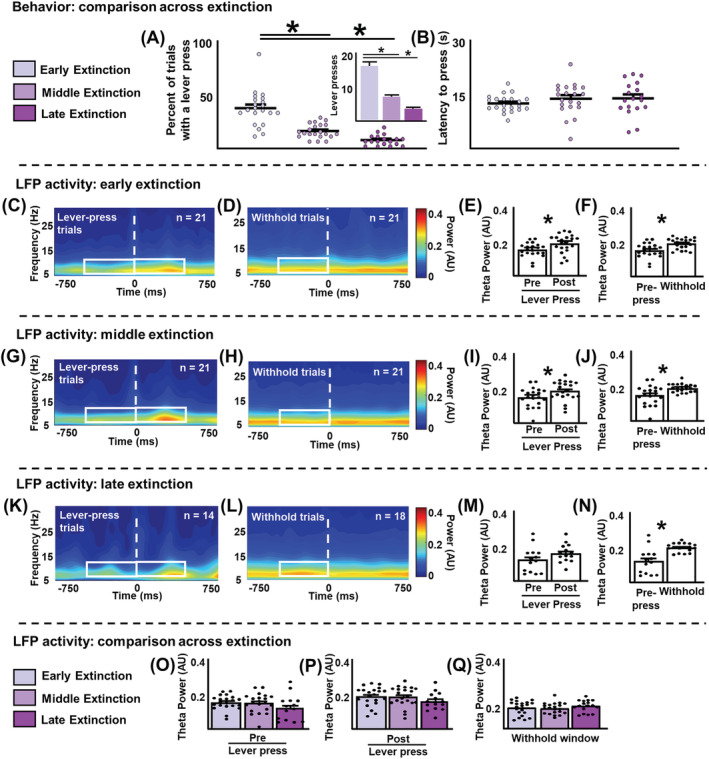
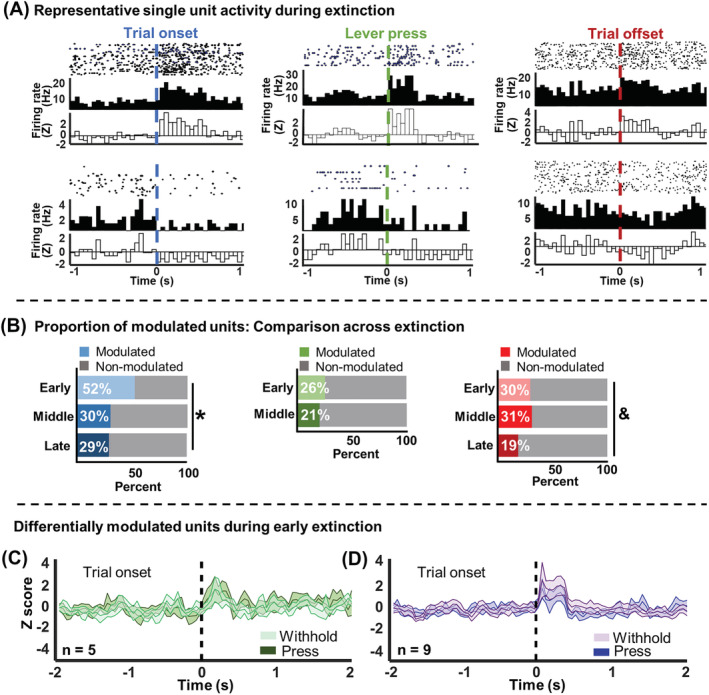
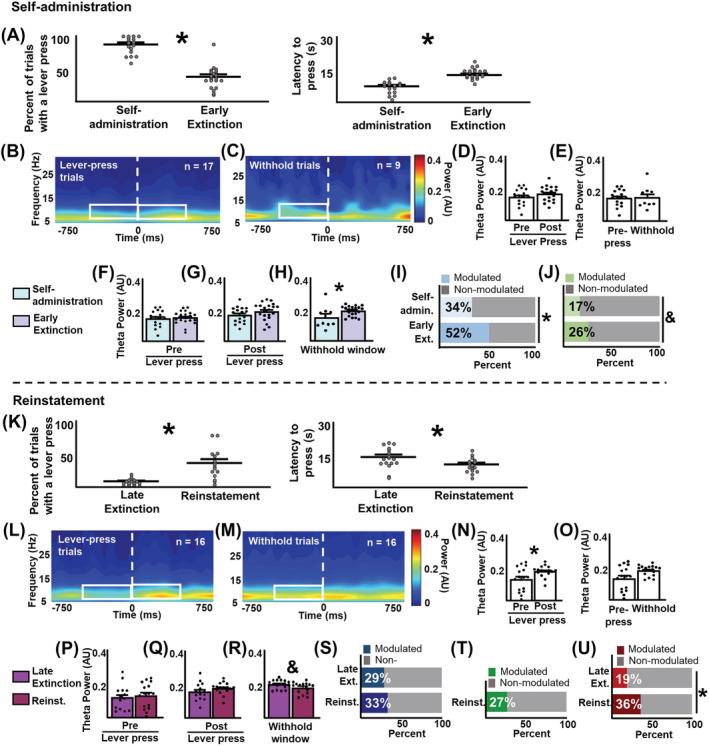
Infralimbic cortical (IL) manipulations indicate that this region mediates extinction learning and suppresses cocaine seeking following cocaine self-administration. However, little work has recorded IL activity during the inhibition of cocaine seeking due to the difficulty of determining precisely when cocaine-seeking behaviour is inhibited within a cocaine-seeking session. The present study used in vivo electrophysiology to examine IL activity across extinction as well as during cocaine self-administration and reinstatement. Sprague-Dawley rats underwent 6-h access cocaine self-administration in which the response lever was available during discrete signalled trials, a procedure which allowed for the comparison between epochs of cocaine seeking versus the inhibition thereof. Subsequently, rats underwent extinction and cocaine-primed reinstatement using the same procedure. Results indicate that theta rhythms (4-10 Hz) dominated IL local-field potential (LFP) activity during all experimental stages. During extinction, theta power fluctuated significantly surrounding the lever press and was lower when rats engaged in cocaine seeking versus when they withheld from doing so. These patterns of oscillatory activity differed from self-administration and reinstatement stages. Single-unit analyses indicate heterogeneity of IL unit responses, supporting the idea that multiple neuronal subpopulations exist within the IL and promote the expression of different and even opposing cocaine-seeking behaviours. Together, these results are consistent with the idea that aggregate synaptic and single-unit activity in the IL represent the engagement of the IL in action monitoring to promote adaptive behaviour in accordance with task contingencies and reveal critical insights into the relationship between IL activity and the inhibition of cocaine seeking.
Keywords: cocaine seeking; cocaine self-administration; in vivo electrophysiology; local-field potential; reinstatement; single units.
© 2021 The Authors. Addiction Biology published by John Wiley & Sons Ltd on behalf of Society for the Study of Addiction.
Conflict of interest statement
Authors have no conflicts of interest to declare.
Figures
Similar articles
-
Extinction of Cocaine Seeking Requires a Window of Infralimbic Pyramidal Neuron Activity after Unreinforced Lever Presses.J Neurosci. 2017 Jun 21;37(25):6075-6086. doi: 10.1523/JNEUROSCI.3821-16.2017. Epub 2017 May 24. J Neurosci. 2017. PMID: 28539416 Free PMC article.
-
Infralimbic Projections to the Nucleus Accumbens Shell and Amygdala Regulate the Encoding of Cocaine Extinction Learning.J Neurosci. 2023 Feb 22;43(8):1348-1359. doi: 10.1523/JNEUROSCI.2023-22.2022. Epub 2023 Jan 19. J Neurosci. 2023. PMID: 36657972 Free PMC article.
-
Vagus Nerve Stimulation (VNS) Modulates Synaptic Plasticity in the Infralimbic Cortex via Trk-B Receptor Activation to Reduce Drug-Seeking in Male Rats.J Neurosci. 2024 Jun 5;44(23):e0107242024. doi: 10.1523/JNEUROSCI.0107-24.2024. J Neurosci. 2024. PMID: 38719446 Free PMC article.
-
Attenuation of cocaine seeking in rats via enhancement of infralimbic cortical activity using stable step-function opsins.Psychopharmacology (Berl). 2019 Jan;236(1):479-490. doi: 10.1007/s00213-018-4964-y. Epub 2018 Jul 12. Psychopharmacology (Berl). 2019. PMID: 30003306 Free PMC article. Review.
-
Infralimbic cortex functioning across motivated behaviors: Can the differences be reconciled?Neurosci Biobehav Rev. 2021 Dec;131:704-721. doi: 10.1016/j.neubiorev.2021.10.002. Epub 2021 Oct 5. Neurosci Biobehav Rev. 2021. PMID: 34624366 Free PMC article. Review.
Cited by
-
Prelimbic cortex and nucleus accumbens core resting state signaling dynamics as a biomarker for cocaine seeking behaviors.Addict Neurosci. 2023 Sep;7:100097. doi: 10.1016/j.addicn.2023.100097. Epub 2023 Apr 19. Addict Neurosci. 2023. PMID: 37396409 Free PMC article.
-
Harnessing the sensing and stimulation function of deep brain-machine interfaces: a new dawn for overcoming substance use disorders.Transl Psychiatry. 2024 Oct 17;14(1):440. doi: 10.1038/s41398-024-03156-8. Transl Psychiatry. 2024. PMID: 39419976 Free PMC article. Review.
-
Prelimbic and infralimbic medial prefrontal cortex neuron activity signals cocaine seeking variables across multiple timescales.Psychopharmacology (Berl). 2023 Mar;240(3):575-594. doi: 10.1007/s00213-022-06287-2. Epub 2022 Dec 5. Psychopharmacology (Berl). 2023. PMID: 36464693 Free PMC article.
-
Perineuronal Nets in the Rat Medial Prefrontal Cortex Alter Hippocampal-Prefrontal Oscillations and Reshape Cocaine Self-Administration Memories.J Neurosci. 2024 Aug 21;44(34):e0468242024. doi: 10.1523/JNEUROSCI.0468-24.2024. J Neurosci. 2024. PMID: 38991791 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
