

Mapping the genetic landscape of DNA double-strand break repair
- PMID: 34672952
- PMCID: PMC9074467
- DOI: 10.1016/j.cell.2021.10.002
Mapping the genetic landscape of DNA double-strand break repair
Abstract
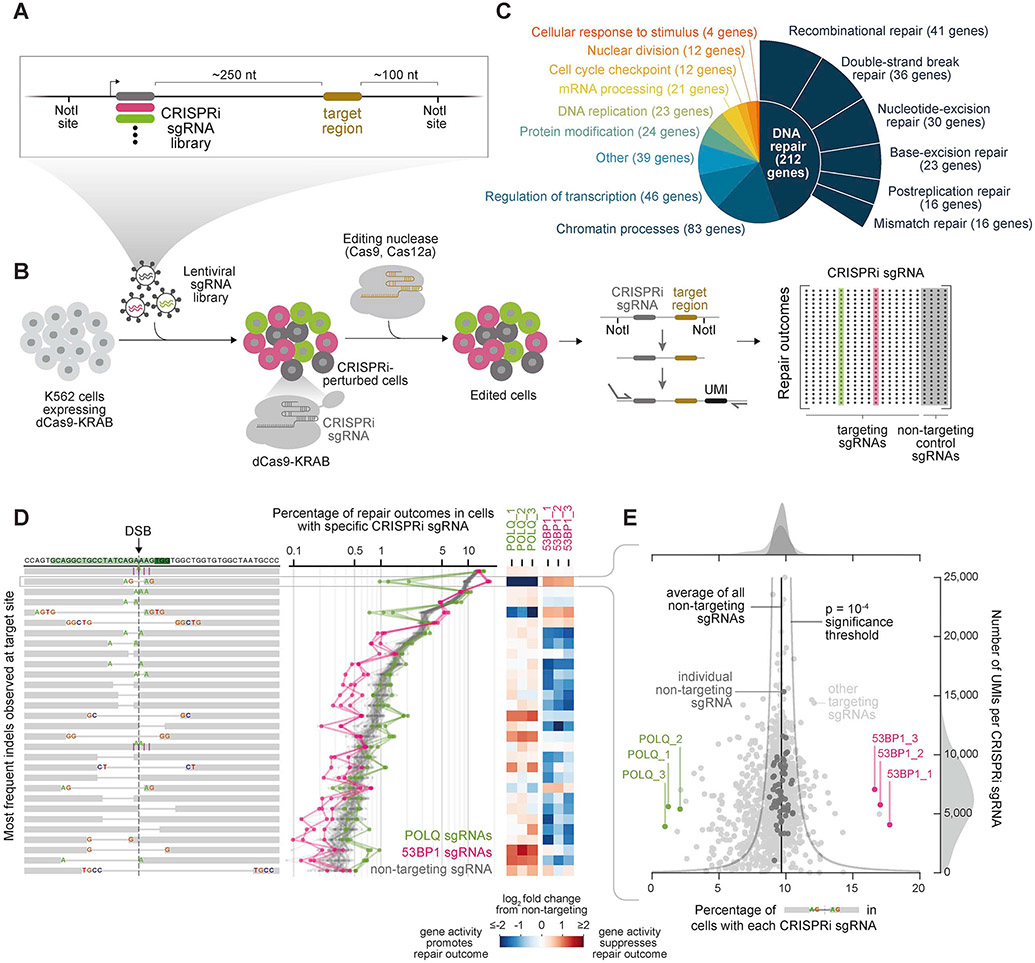
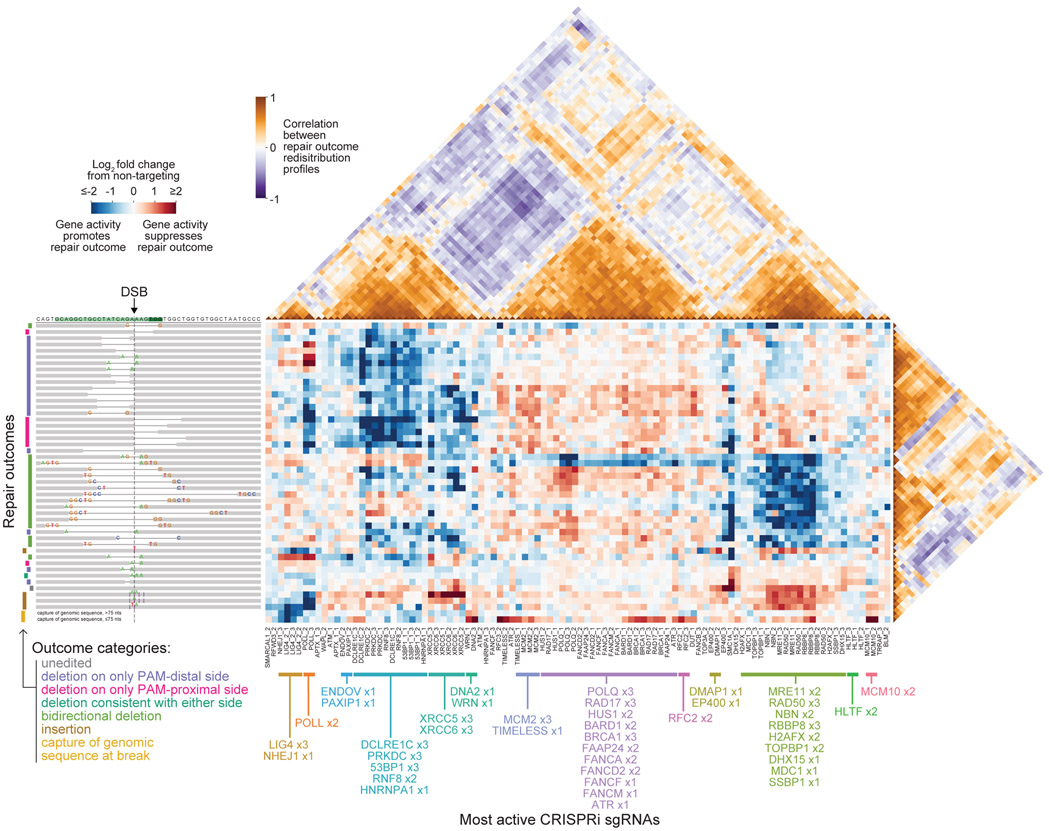
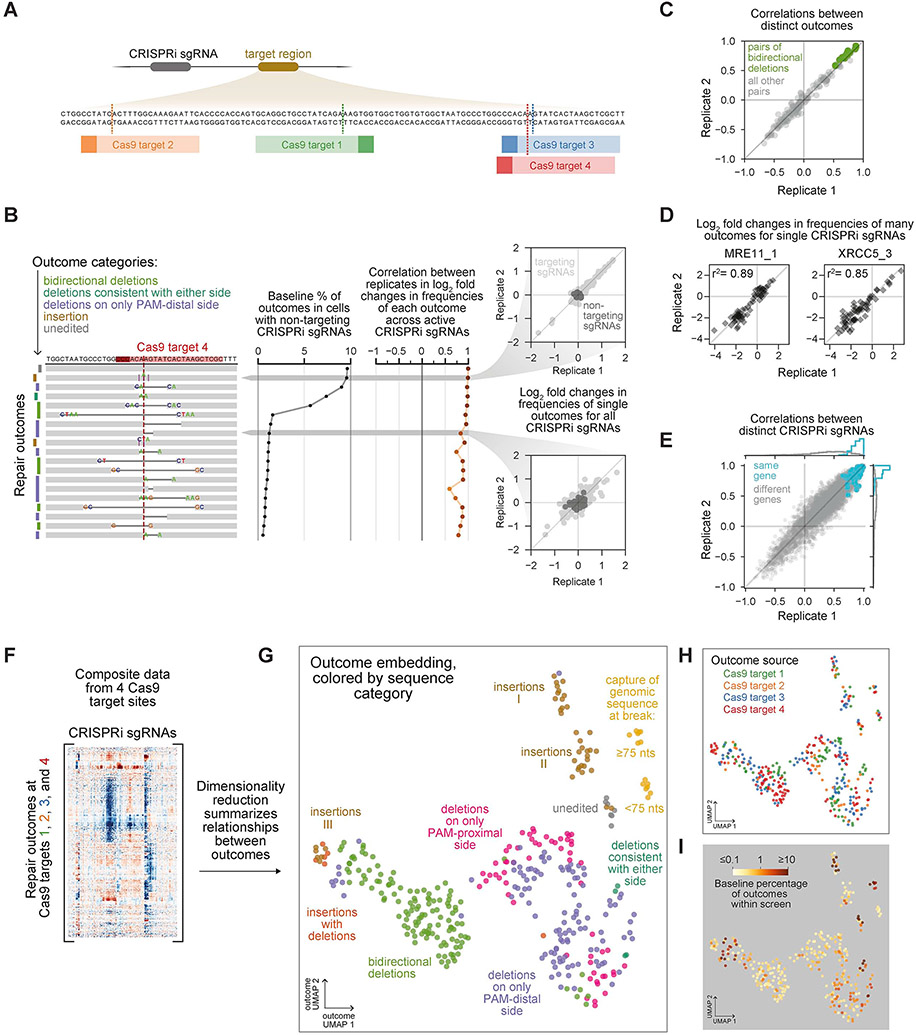
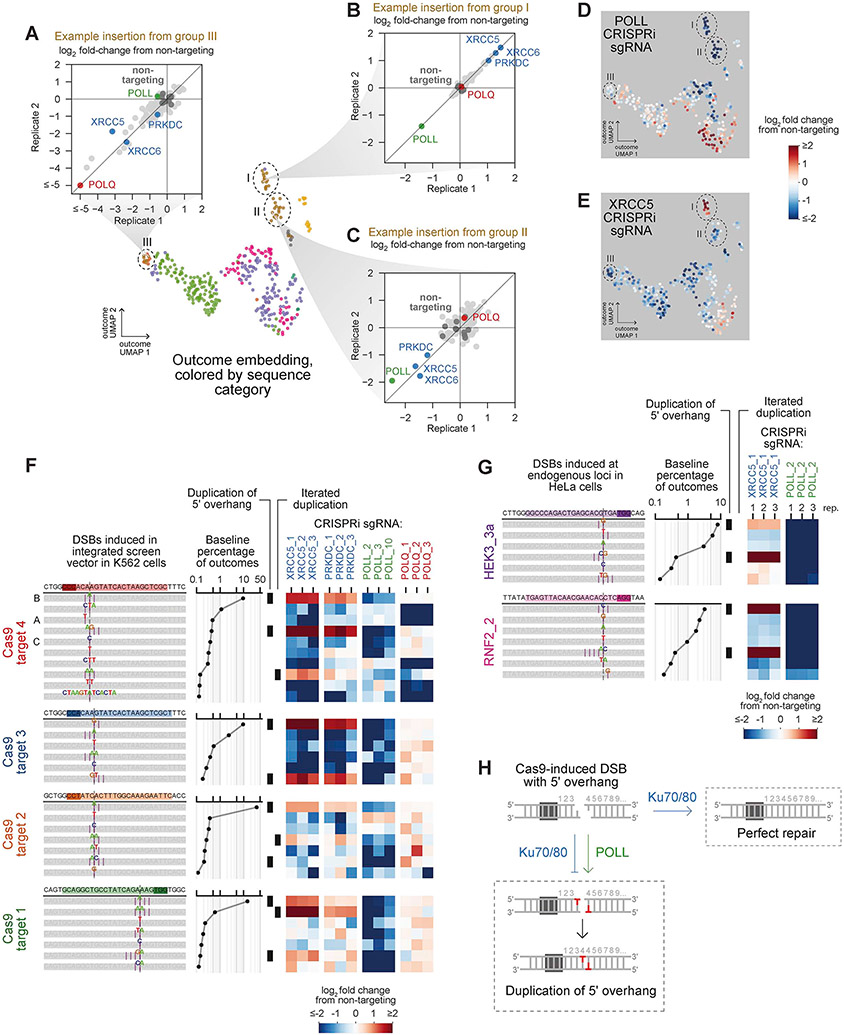
Cells repair DNA double-strand breaks (DSBs) through a complex set of pathways critical for maintaining genomic integrity. To systematically map these pathways, we developed a high-throughput screening approach called Repair-seq that measures the effects of thousands of genetic perturbations on mutations introduced at targeted DNA lesions. Using Repair-seq, we profiled DSB repair products induced by two programmable nucleases (Cas9 and Cas12a) in the presence or absence of oligonucleotides for homology-directed repair (HDR) after knockdown of 476 genes involved in DSB repair or associated processes. The resulting data enabled principled, data-driven inference of DSB end joining and HDR pathways. Systematic interrogation of this data uncovered unexpected relationships among DSB repair genes and demonstrated that repair outcomes with superficially similar sequence architectures can have markedly different genetic dependencies. This work provides a foundation for mapping DNA repair pathways and for optimizing genome editing across diverse modalities.
Keywords: CRISPR-Cas9; DNA repair; double-strand breaks; functional genomics; genome editing.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests Editas Medicine was involved in this work and provided reagents. B.A. was a member of a ThinkLab Advisory Board for, and holds equity in, Celsius Therapeutics. J.A.H. is a consultant for Tessera Therapeutics. J.S.W. declares outside interest in 5 AM Venture, Amgen, Chroma Medicine, KSQ Therapeutics, Maze Therapeutics, Tenaya Therapeutics, Tessera Therapeutics, and Third Rock Ventures. A.B. and C.C.-R. are former employees and shareholders of Editas Medicine and were employed by Editas at the time this work was conducted. J.A.H. and B.A. have filed patent applications on related work.
Figures
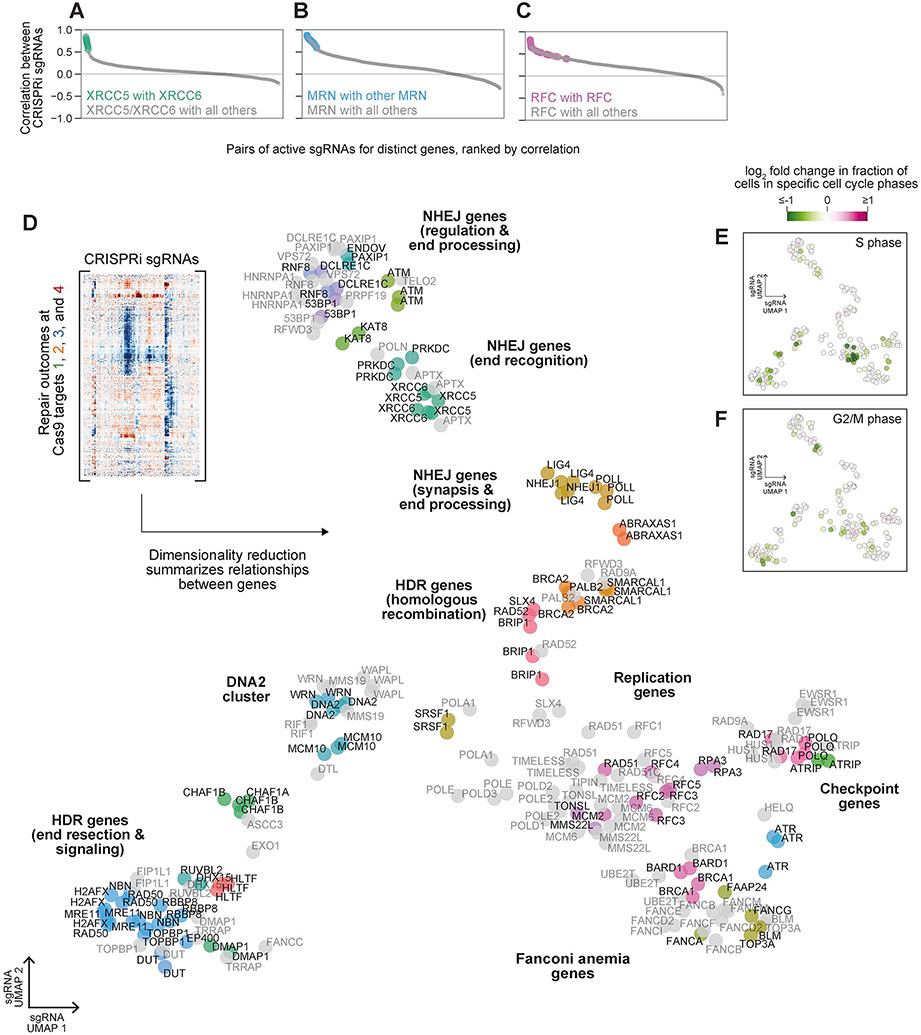
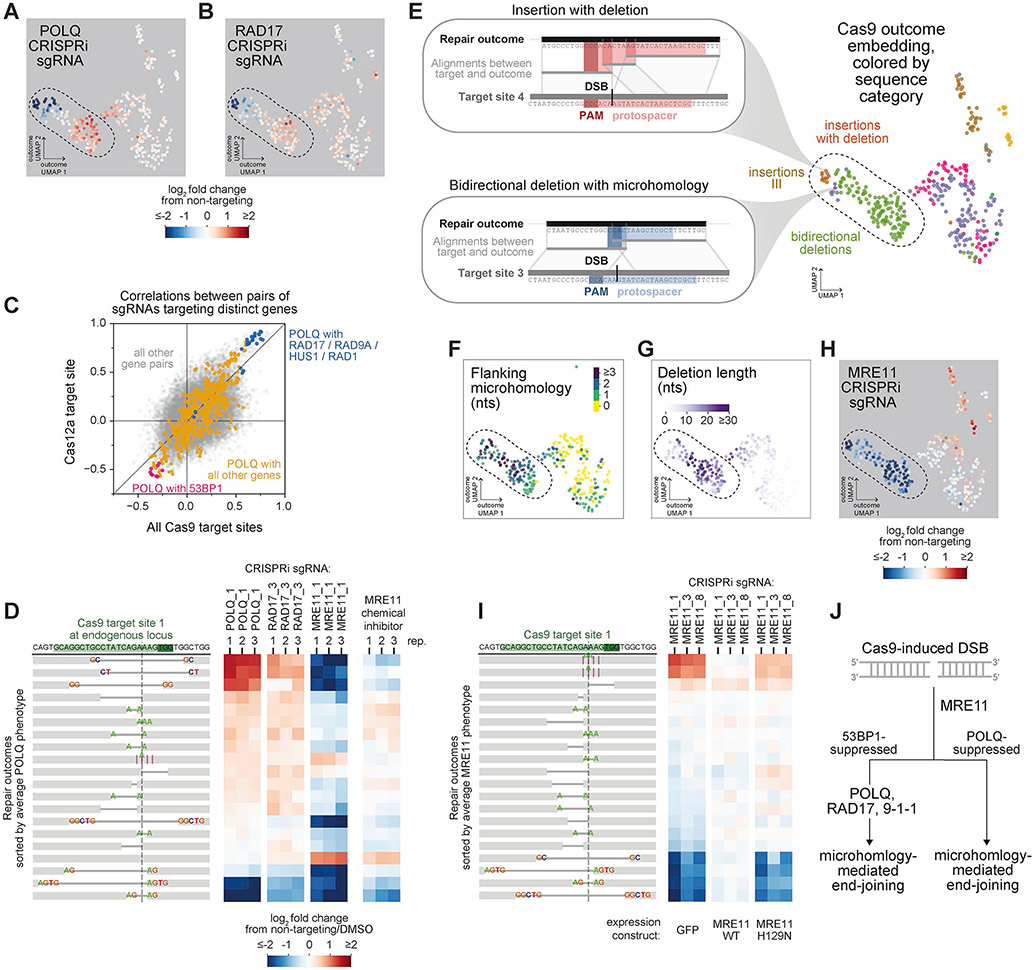
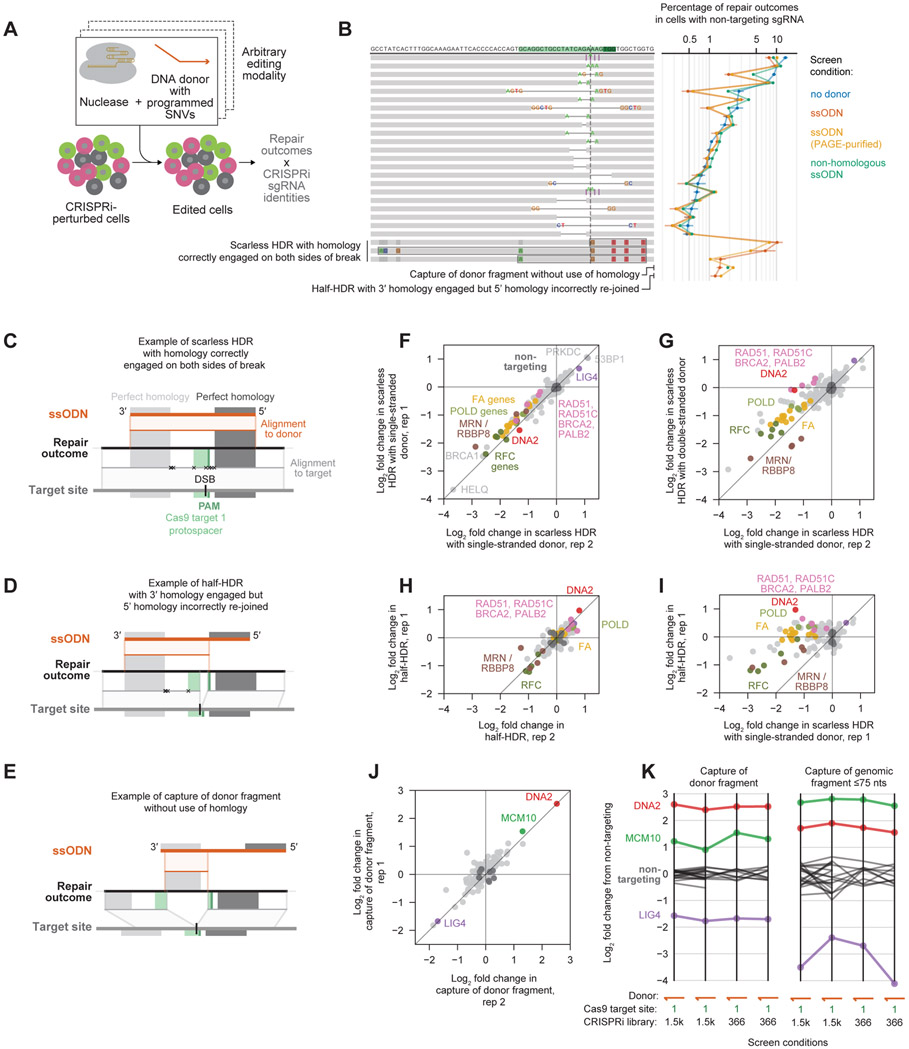
Comment in
-
Illuminating the path to DNA repair.Cell. 2021 Oct 28;184(22):5503-5505. doi: 10.1016/j.cell.2021.10.005. Cell. 2021. PMID: 34715020
-
Profiling double-strand break repair.Nat Methods. 2021 Dec;18(12):1449. doi: 10.1038/s41592-021-01352-8. Nat Methods. 2021. PMID: 34862500 No abstract available.
-
Improving CRISPR tools by elucidating DNA repair.Nat Biotechnol. 2021 Dec;39(12):1512-1514. doi: 10.1038/s41587-021-01149-2. Nat Biotechnol. 2021. PMID: 34873327 No abstract available.
-
Repair-seq: Seeking and Perturbing DNA Repair.CRISPR J. 2021 Dec;4(6):773-775. doi: 10.1089/crispr.2021.29140.dme. CRISPR J. 2021. PMID: 34935489 No abstract available.
References
-
- Anzalone AV, Koblan LW, and Liu DR (2020). Genome editing with CRISPR–Cas nucleases, base editors, transposases and prime editors. Nat. Biotechnol - PubMed
-
- Bothmer A, Bothmer A, Gareau KW, Abdulkerim HS, Abdulkerim HS, Buquicchio F, Buquicchio F, Buquicchio F, Cohen L, Cohen L, et al. (2020). Detection and Modulation of DNA Translocations during Multi-Gene Genome Editing in T Cells. Cris. J 3, 177–187. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
