

A single amino acid mutation in the mouse MEIG1 protein disrupts a cargo transport system necessary for sperm formation
- PMID: 34673028
- PMCID: PMC8592874
- DOI: 10.1016/j.jbc.2021.101312
A single amino acid mutation in the mouse MEIG1 protein disrupts a cargo transport system necessary for sperm formation
Abstract
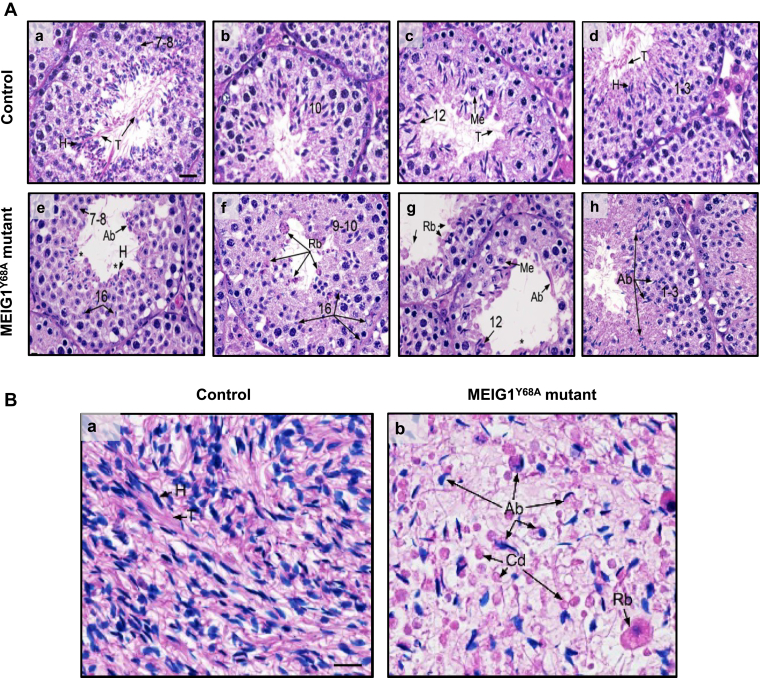
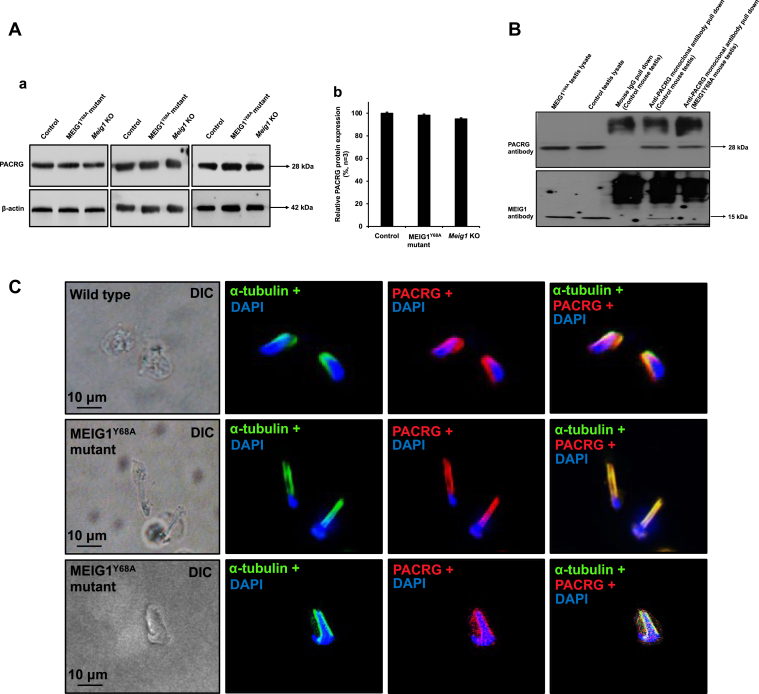
Mammalian spermatogenesis is a highly coordinated process that requires cooperation between specific proteins to coordinate diverse biological functions. For example, mouse Parkin coregulated gene (PACRG) recruits meiosis-expressed gene 1 (MEIG1) to the manchette during normal spermiogenesis. Here we mutated Y68 of MEIG1 using the CRISPR/cas9 system and examined the biological and physiological consequences in mice. All homozygous mutant males examined were completely infertile, and sperm count was dramatically reduced. The few developed sperm were immotile and displayed multiple abnormalities. Histological staining showed impaired spermiogenesis in these mutant mice. Immunofluorescent staining further revealed that this mutant MEIG1 was still present in the cell body of spermatocytes, but also that more MEIG1 accumulated in the acrosome region of round spermatids. The mutant MEIG1 and a cargo protein of the MEIG1/PACRG complex, sperm-associated antigen 16L (SPAG16L), were no longer found to be present in the manchette; however, localization of the PACRG component was not changed in the mutants. These findings demonstrate that Y68 of MEIG1 is a key amino acid required for PACRG to recruit MEIG1 to the manchette to transport cargo proteins during sperm flagella formation. Given that MEIG1 and PACRG are conserved in humans, small molecules that block MEIG1/PACRG interaction are likely ideal targets for the development of male contraconception drugs.
Keywords: MEIG1; PACRG; SPAG16; cargo transport; male fertility; manchette; single amino acid mutation; spermiogenesis.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures





References
-
- Don J., Wolgemuth D.J. Identification and characterization of the regulated pattern of expression of a novel mouse gene, meg1, during the meiotic cell cycle. Cell Growth Differ. 1992;3:495–505. - PubMed
-
- Salzberg Y., Eldar T., Karminsky O.D., Itach S.B., Pietrokovski S., Don J. Meig1 deficiency causes a severe defect in mouse spermatogenesis. Dev. Biol. 2010;338:158–167. - PubMed
-
- Don J., Winer M.A., Wolgemuth D.J. Developmentally regulated expression during gametogenesis of the murine gene meg1 suggests a role in meiosis. Mol. Reprod. Dev. 1994;38:16–23. - PubMed
-
- Chen-Moses A., Malkov M., Shalom S., Ever L., Don J. A switch in the phosphorylation state of the dimeric form of the Meg1 protein correlates with progression through meiosis in the mouse. Cell Growth Differ. 1997;8:711–719. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
