

T cell receptor-targeted immunotherapeutics drive selective in vivo HIV- and CMV-specific T cell expansion in humanized mice
- PMID: 34673568
- PMCID: PMC8631598
- DOI: 10.1172/JCI141051
T cell receptor-targeted immunotherapeutics drive selective in vivo HIV- and CMV-specific T cell expansion in humanized mice
Abstract
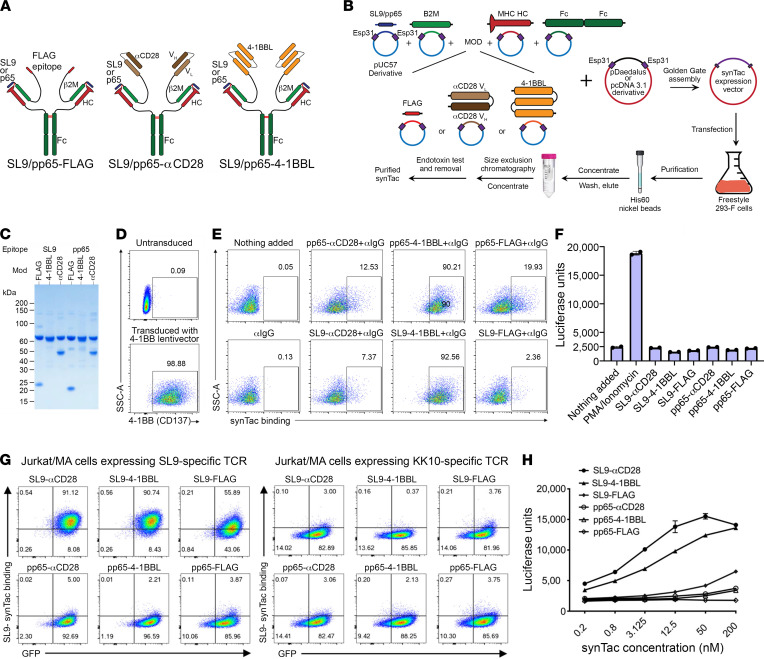
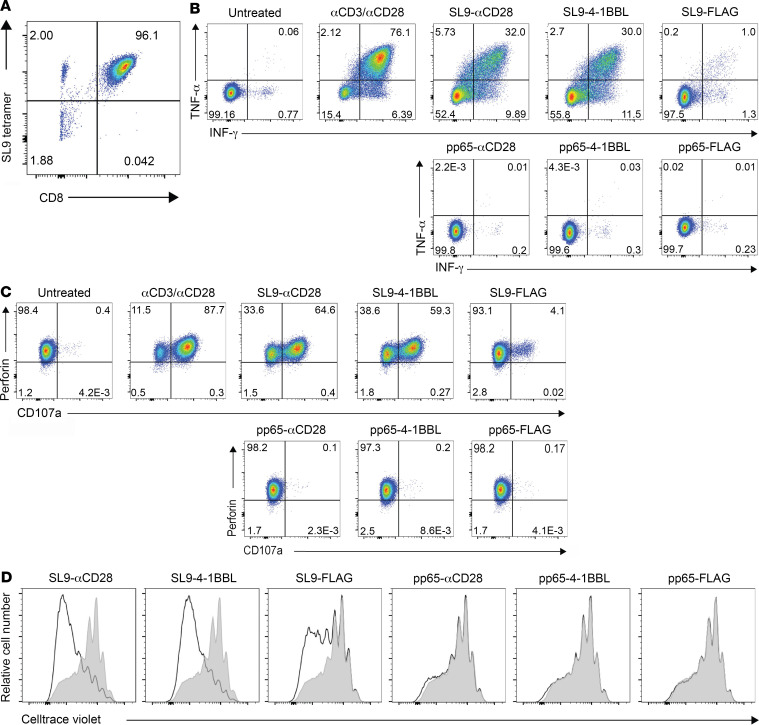
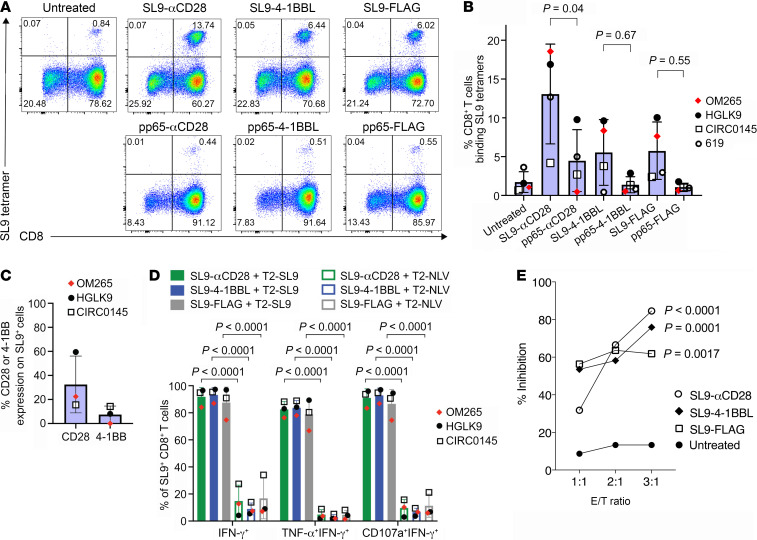
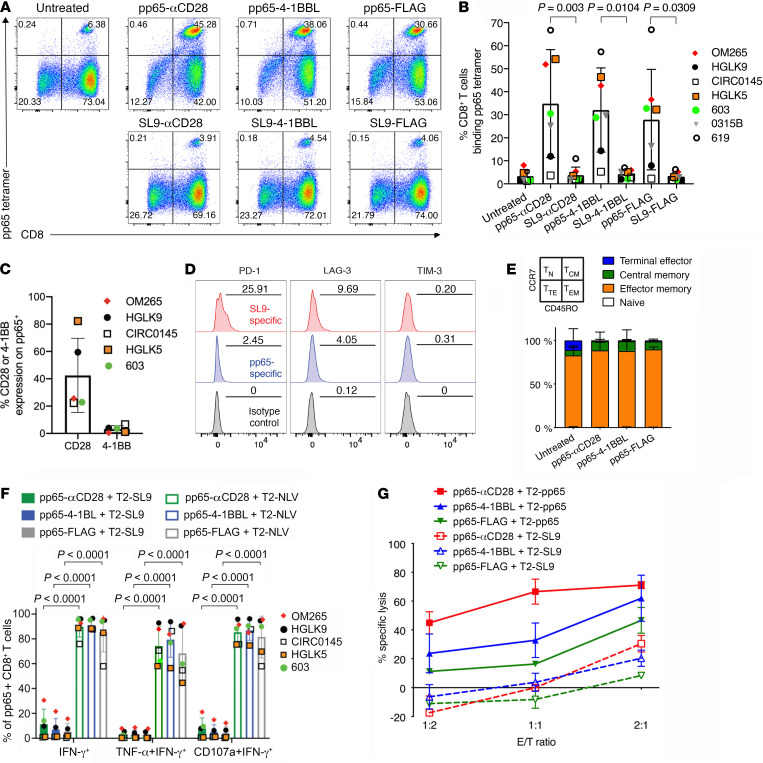
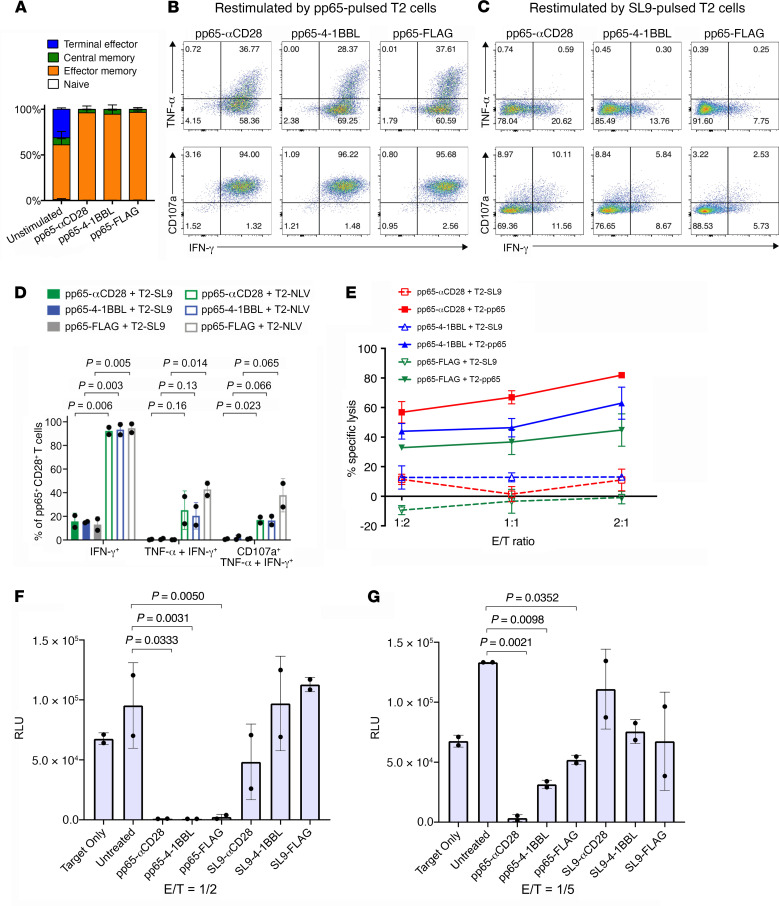
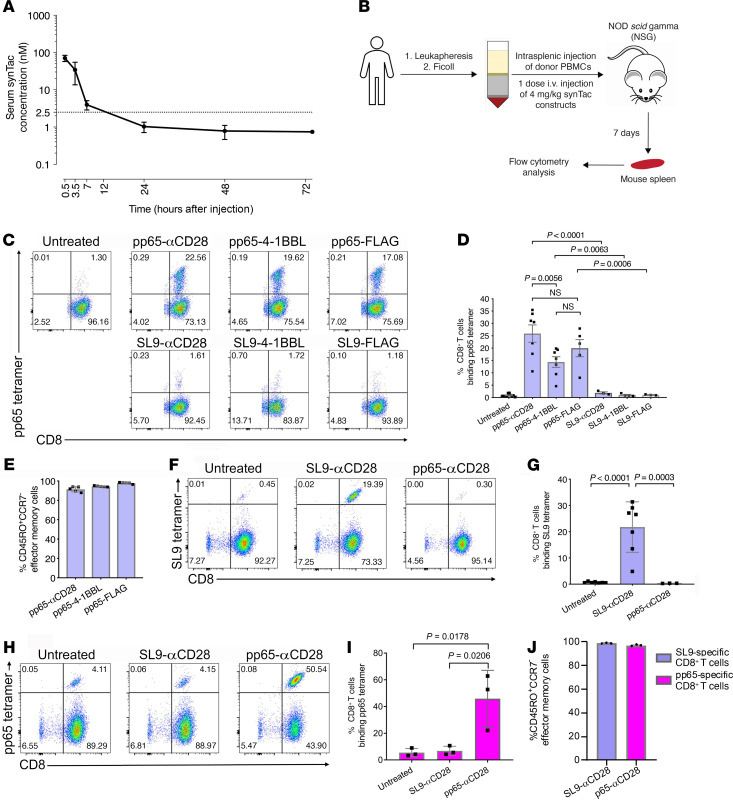
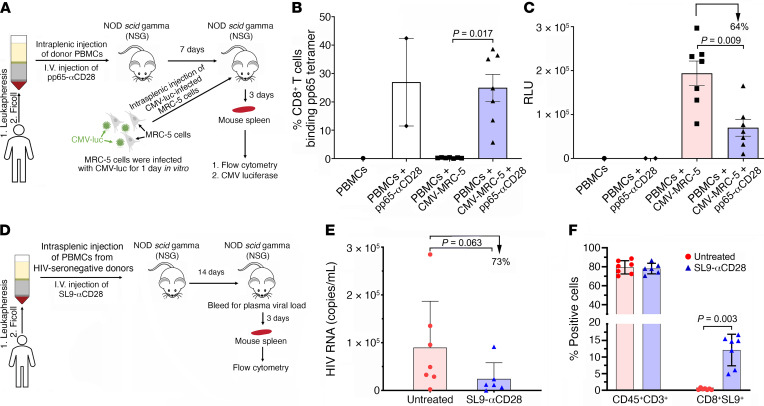
To delineate the in vivo role of different costimulatory signals in activating and expanding highly functional virus-specific cytotoxic CD8+ T cells, we designed synTacs, infusible biologics that recapitulate antigen-specific T cell activation signals delivered by antigen-presenting cells. We constructed synTacs consisting of dimeric Fc-domain scaffolds linking CD28- or 4-1BB-specific ligands to HLA-A2 MHC molecules covalently tethered to HIV- or CMV-derived peptides. Treatment of HIV-infected donor PBMCs with synTacs bearing HIV- or CMV-derived peptides induced vigorous and selective ex vivo expansion of highly functional HIV- and/or CMV-specific CD8+ T cells, respectively, with potent antiviral activities. Intravenous injection of HIV- or CMV-specific synTacs into immunodeficient mice intrasplenically engrafted with donor PBMCs markedly and selectively expanded HIV-specific (32-fold) or CMV-specific (46-fold) human CD8+ T cells populating their spleens. Notably, these expanded HIV- or CMV-specific CD8+ T cells directed potent in vivo suppression of HIV or CMV infections in the humanized mice, providing strong rationale for consideration of synTac-based approaches as a therapeutic strategy to cure HIV and treat CMV and other viral infections. The synTac platform flexibility supports facile delineation of in vivo effects of different costimulatory signals on patient-derived virus-specific CD8+ T cells, enabling optimization of individualized therapies, including HIV cure strategies.
Keywords: AIDS/HIV; Costimulation; Immunology; T cells.
Conflict of interest statement
Figures

Similar articles
-
Refining human T-cell immunotherapy of cytomegalovirus disease: a mouse model with 'humanized' antigen presentation as a new preclinical study tool.Med Microbiol Immunol. 2016 Dec;205(6):549-561. doi: 10.1007/s00430-016-0471-0. Epub 2016 Aug 18. Med Microbiol Immunol. 2016. PMID: 27539576 Review.
-
Cytomegalovirus-Mediated T Cell Receptor Repertoire Perturbation Is Present in Early Life.Front Immunol. 2020 Sep 30;11:1587. doi: 10.3389/fimmu.2020.01587. eCollection 2020. Front Immunol. 2020. PMID: 33101265 Free PMC article.
-
Expansion of human cytomegalovirus-specific T lymphocytes from unfractionated peripheral blood mononuclear cells with artificial antigen-presenting cells.Transfusion. 2007 Nov;47(11):2143-52. doi: 10.1111/j.1537-2995.2007.01439.x. Transfusion. 2007. PMID: 17958544
-
CMV-infected kidney grafts drive the expansion of blood-borne CMV-specific T cells restricted by shared class I HLA molecules via presentation on donor cells.Am J Transplant. 2018 Aug;18(8):1904-1913. doi: 10.1111/ajt.14672. Epub 2018 Mar 3. Am J Transplant. 2018. PMID: 29377506
-
Gene transfer for activation of CMV specific T cells.Hum Immunol. 2004 May;65(5):565-70. doi: 10.1016/j.humimm.2004.02.015. Hum Immunol. 2004. PMID: 15172458 Review.
Cited by
-
Agonism of 4-1BB for immune therapy: a perspective on possibilities and complications.Front Immunol. 2023 Aug 17;14:1228486. doi: 10.3389/fimmu.2023.1228486. eCollection 2023. Front Immunol. 2023. PMID: 37662949 Free PMC article. Review.
-
Memory inflation: Beyond the acute phase of viral infection.Cell Prolif. 2024 Dec;57(12):e13705. doi: 10.1111/cpr.13705. Epub 2024 Jul 11. Cell Prolif. 2024. PMID: 38992867 Free PMC article. Review.
-
Development of a Stable Peptide-Major Histocompatibility Complex (MHC) via Sortase and Click Chemistry.ACS Pharmacol Transl Sci. 2024 May 24;7(6):1746-1757. doi: 10.1021/acsptsci.3c00268. eCollection 2024 Jun 14. ACS Pharmacol Transl Sci. 2024. PMID: 38898944 Free PMC article.
-
Recent advances on anti-HIV chimeric antigen receptor-T-cell treatment to provide sustained HIV remission.Curr Opin HIV AIDS. 2024 Jul 1;19(4):169-178. doi: 10.1097/COH.0000000000000858. Epub 2024 May 1. Curr Opin HIV AIDS. 2024. PMID: 38695148 Free PMC article. Review.
-
IL-2-armored peptide-major histocompatibility class I bispecific antibodies redirect antiviral effector memory CD8+ T cells to induce potent anti-cancer cytotoxic activity with limited cytokine release.MAbs. 2024 Jan-Dec;16(1):2395499. doi: 10.1080/19420862.2024.2395499. Epub 2024 Aug 28. MAbs. 2024. PMID: 39205483 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
- R01 CA198095/CA/NCI NIH HHS/United States
- 75N93022D00005/AI/NIAID NIH HHS/United States
- 75N99020D00005/OF/ORFDO NIH HHS/United States
- R01 DA044584/DA/NIDA NIH HHS/United States
- U54 GM094662/GM/NIGMS NIH HHS/United States
- 75N95020D00005/DA/NIDA NIH HHS/United States
- R01 AI145024/AI/NIAID NIH HHS/United States
- R01 DA048609/DA/NIDA NIH HHS/United States
- P30 AI124414/AI/NIAID NIH HHS/United States
- U01 GM094665/GM/NIGMS NIH HHS/United States
- P30 CA013330/CA/NCI NIH HHS/United States
- UM1 AI126617/AI/NIAID NIH HHS/United States
