

The Plasmodium NOT1-G paralogue is an essential regulator of sexual stage maturation and parasite transmission
- PMID: 34673764
- PMCID: PMC8562791
- DOI: 10.1371/journal.pbio.3001434
The Plasmodium NOT1-G paralogue is an essential regulator of sexual stage maturation and parasite transmission
Abstract
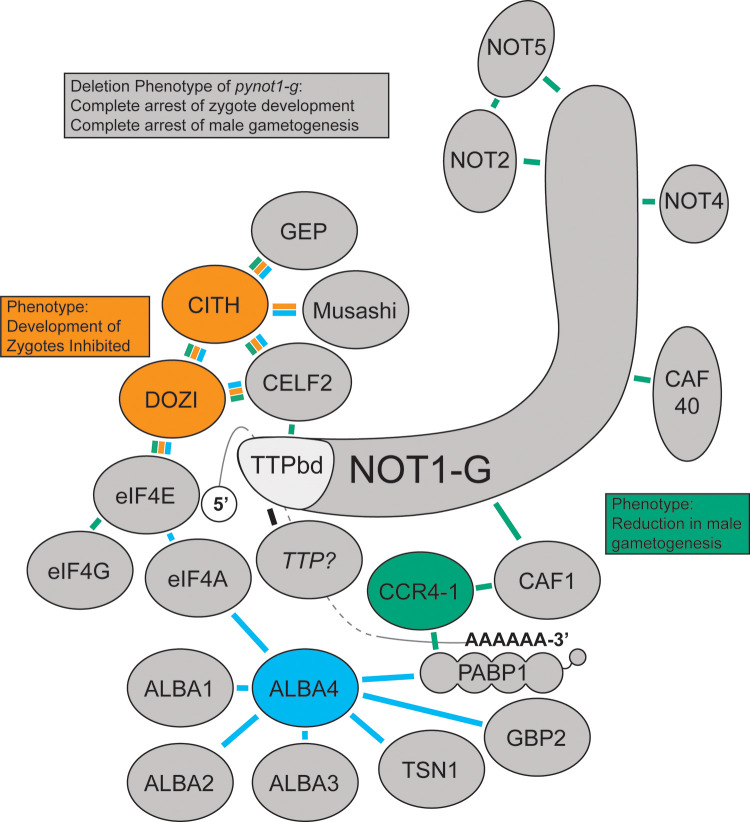
Productive transmission of malaria parasites hinges upon the execution of key transcriptional and posttranscriptional regulatory events. While much is now known about how specific transcription factors activate or repress sexual commitment programs, far less is known about the production of a preferred mRNA homeostasis following commitment and through the host-to-vector transmission event. Here, we show that in Plasmodium parasites, the NOT1 scaffold protein of the CAF1/CCR4/Not complex is duplicated, and one paralogue is dedicated for essential transmission functions. Moreover, this NOT1-G paralogue is central to the sex-specific functions previously associated with its interacting partners, as deletion of not1-g in Plasmodium yoelii leads to a comparable or complete arrest phenotype for both male and female parasites. We show that, consistent with its role in other eukaryotes, PyNOT1-G localizes to cytosolic puncta throughout much of the Plasmodium life cycle. PyNOT1-G is essential to both the complete maturation of male gametes and to the continued development of the fertilized zygote originating from female parasites. Comparative transcriptomics of wild-type and pynot1-g- parasites shows that loss of PyNOT1-G leads to transcript dysregulation preceding and during gametocytogenesis and shows that PyNOT1-G acts to preserve mRNAs that are critical to sexual and early mosquito stage development. Finally, we demonstrate that the tristetraprolin (TTP)-binding domain, which acts as the typical organization platform for RNA decay (TTP) and RNA preservation (ELAV/HuR) factors is dispensable for PyNOT1-G's essential blood stage functions but impacts host-to-vector transmission. Together, we conclude that a NOT1-G paralogue in Plasmodium fulfills the complex transmission requirements of both male and female parasites.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures






References
-
- World Health Organization. World Malaria Report 2020. Available from: https://www.who.int/publications/i/item/9789240015791.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
