

Serotonin modulates melatonin synthesis as an autocrine neurotransmitter in the pineal gland
- PMID: 34675083
- PMCID: PMC8639368
- DOI: 10.1073/pnas.2113852118
Serotonin modulates melatonin synthesis as an autocrine neurotransmitter in the pineal gland
Abstract
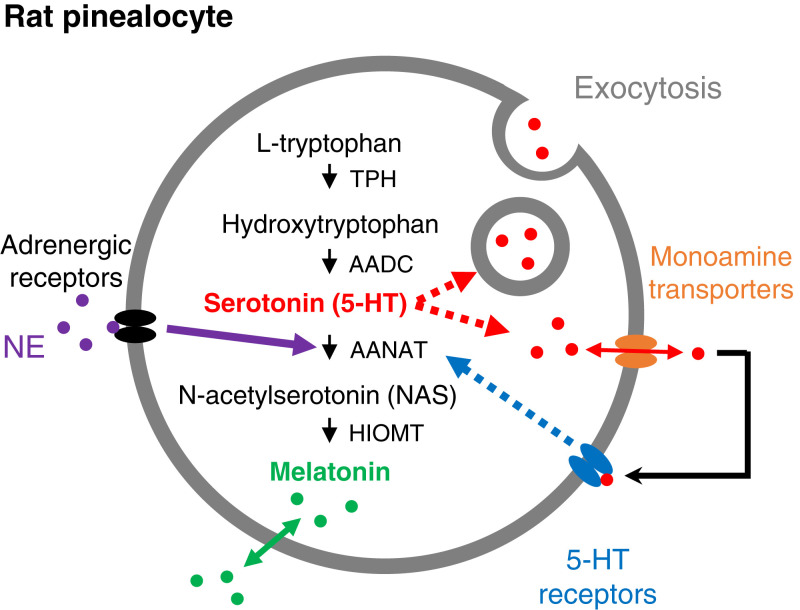
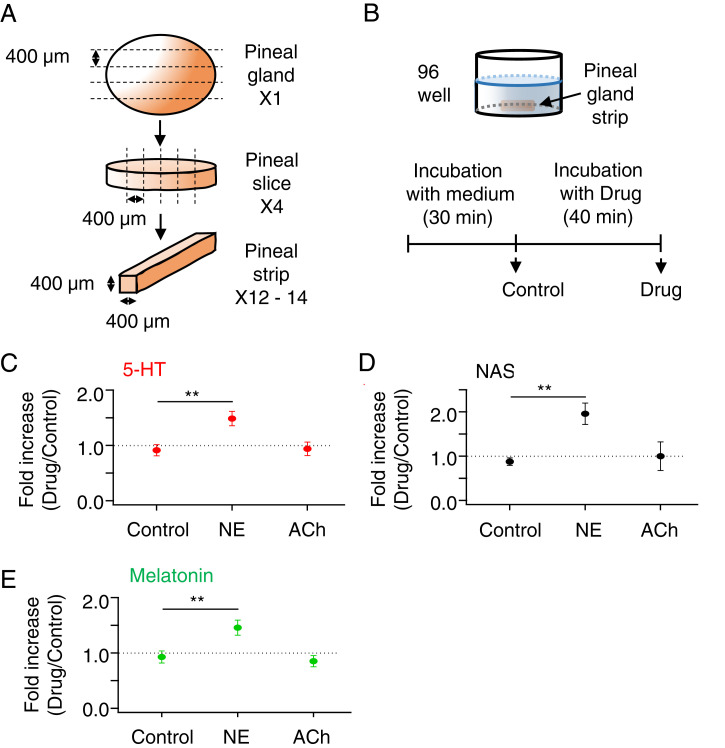
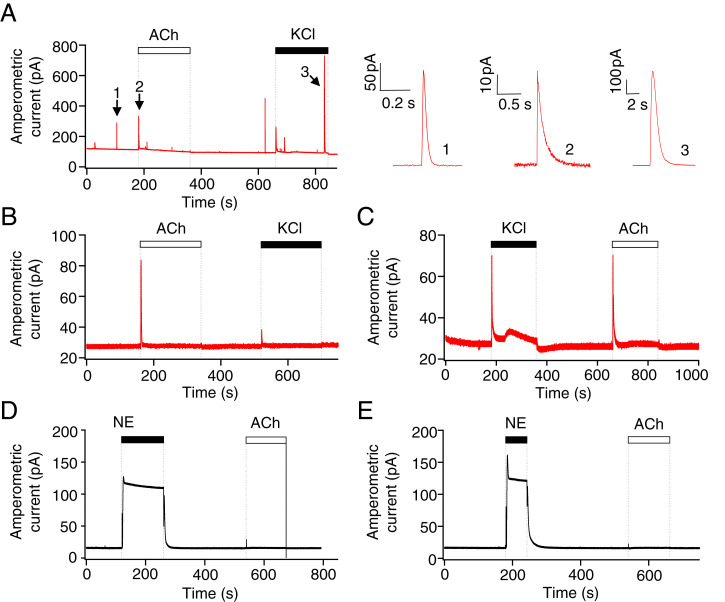
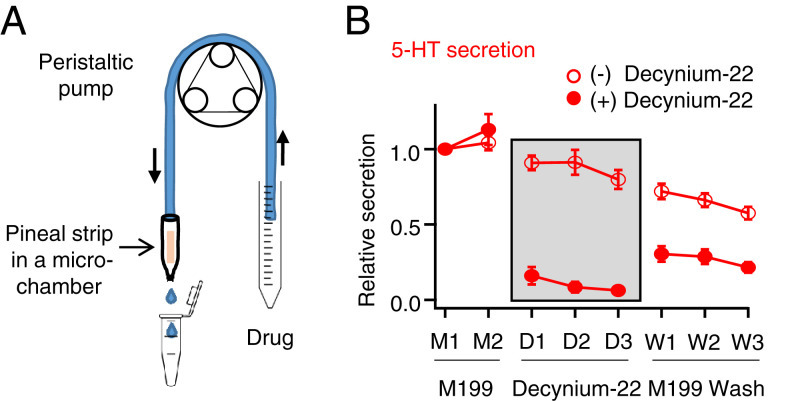
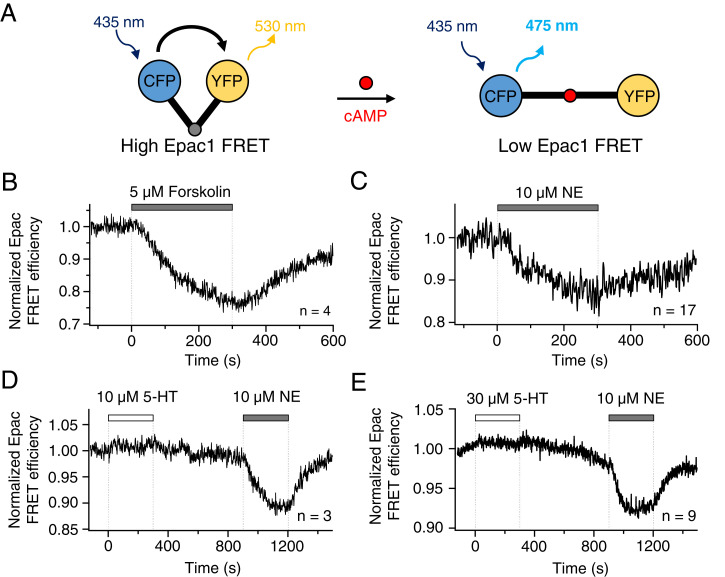
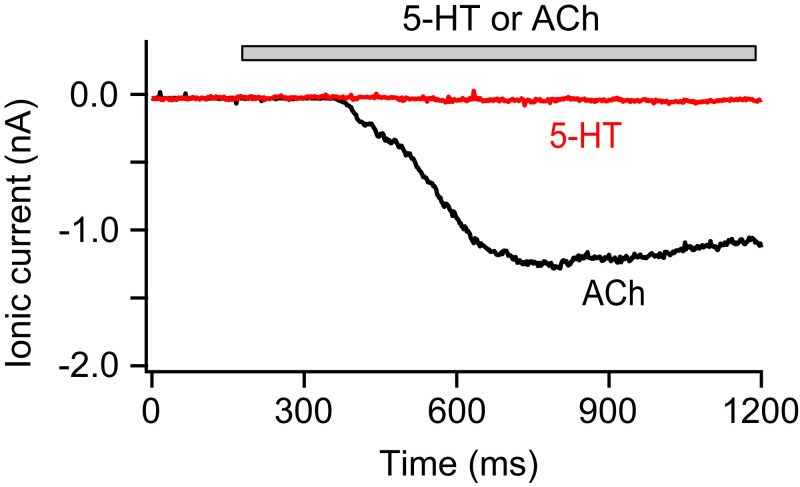
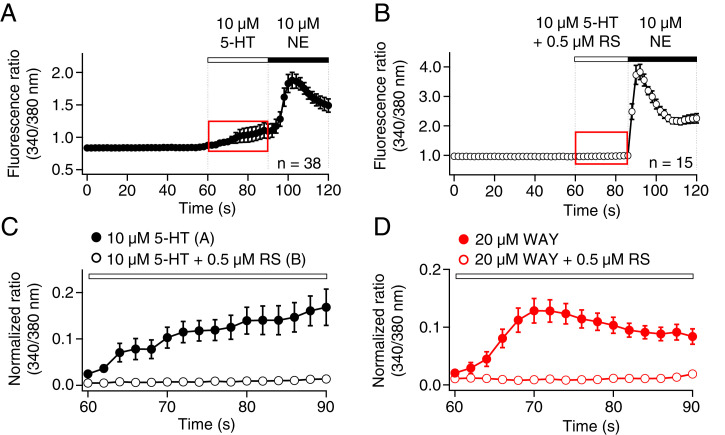
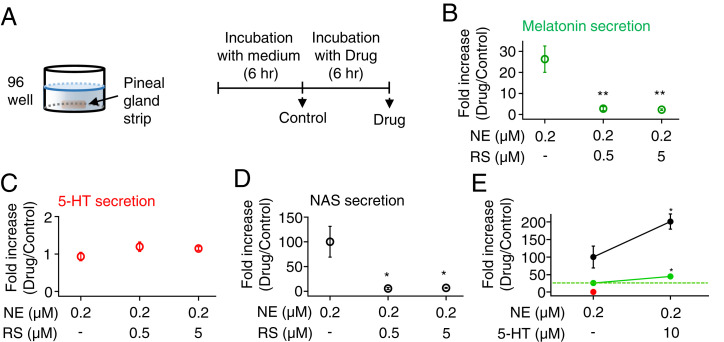
The pineal gland secretes melatonin principally at night. Regulated by norepinephrine released from sympathetic nerve terminals, adrenergic receptors on pinealocytes activate aralkylamine N-acetyltransferase that converts 5-hydroxytryptamine (5-HT, serotonin) to N-acetylserotonin, the precursor of melatonin. Previous studies from our group and others reveal significant constitutive secretion of 5-HT from pinealocytes. Here, using mass spectrometry, we demonstrated that the 5-HT is secreted primarily via a decynium-22-sensitive equilibrative plasma membrane monoamine transporter instead of by typical exocytotic quantal secretion. Activation of the endogenous 5-HT receptors on pinealocytes evoked an intracellular Ca2+ rise that was blocked by RS-102221, an antagonist of 5-HT2C receptors. Applied 5-HT did not evoke melatonin secretion by itself, but it did potentiate melatonin secretion evoked by submaximal norepinephrine. In addition, RS-102221 reduced the norepinephrine-induced melatonin secretion in strips of pineal gland, even when no exogenous 5-HT was added, suggesting that the 5-HT that is constitutively released from pinealocytes accumulates enough in the tissue to act as an autocrine feedback signal sensitizing melatonin release.
Keywords: N-acetylserotonin; melatonin; pineal gland; plasma membrane monoamine transporter; serotonin receptor.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Klein D. C., et al., The melatonin rhythm-generating enzyme: Molecular regulation of serotonin N-acetyltransferase in the pineal gland. Recent Prog. Horm. Res. 52, 307–357, discussion 357–358 (1997). - PubMed
-
- Sun X., Liu T., Deng J., Borjigin J., Long-term in vivo pineal microdialysis. J. Pineal Res. 35, 118–124 (2003). - PubMed
-
- Møller M., Baeres F. M., The anatomy and innervation of the mammalian pineal gland. Cell Tissue Res. 309, 139–150 (2002). - PubMed
-
- Masson J., Emerit M. B., Hamon M., Darmon M., Serotonergic signaling: Multiple effectors and pleiotropic effects. Wiley Interdiscip. Rev. Membr. Transp. Signal. 1, 685–713 (2012).
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
