

Immunogenicity of In Vitro-Transcribed RNA
- PMID: 34677064
- PMCID: PMC9127547
- DOI: 10.1021/acs.accounts.1c00521
Immunogenicity of In Vitro-Transcribed RNA
Abstract
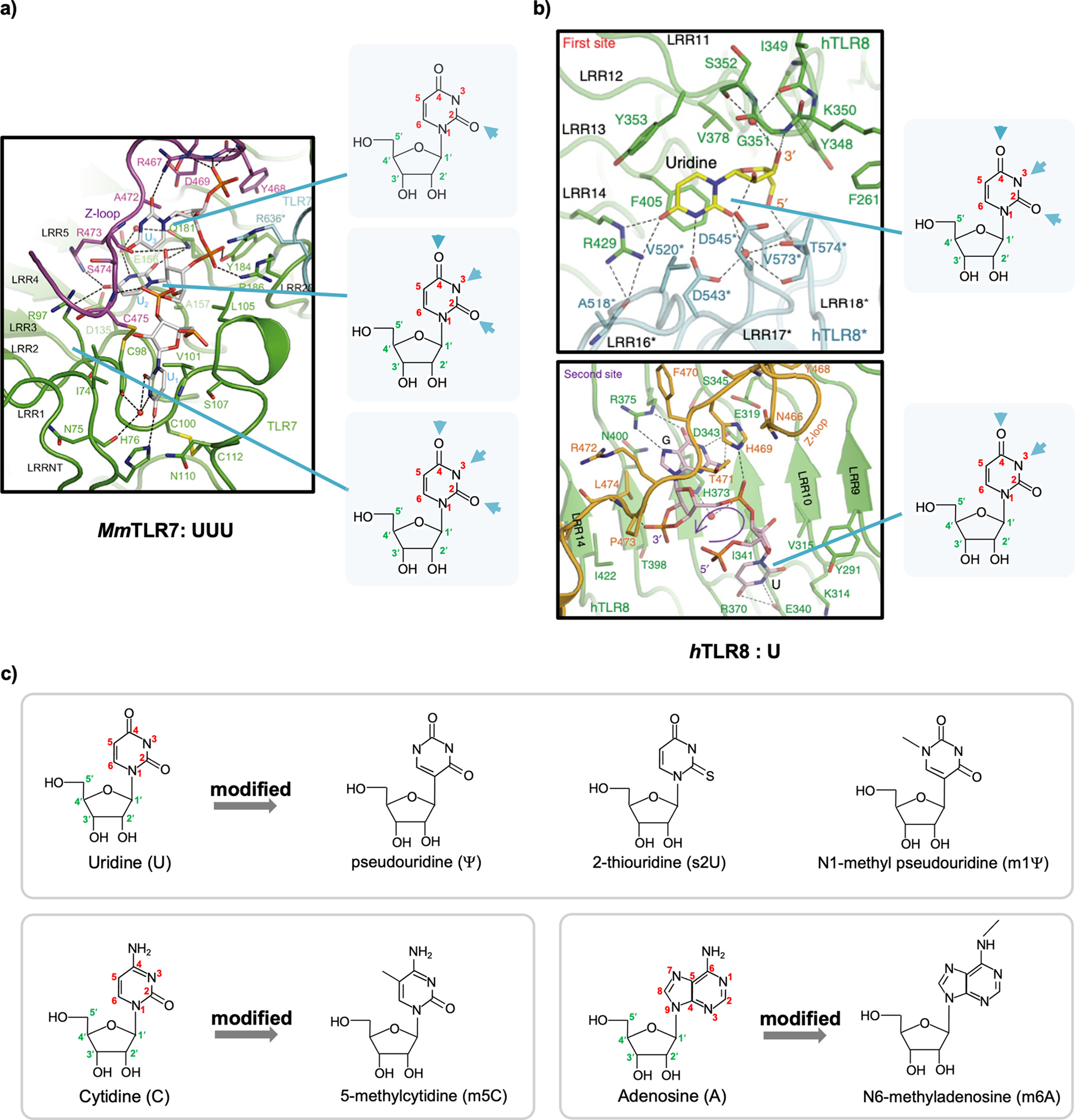
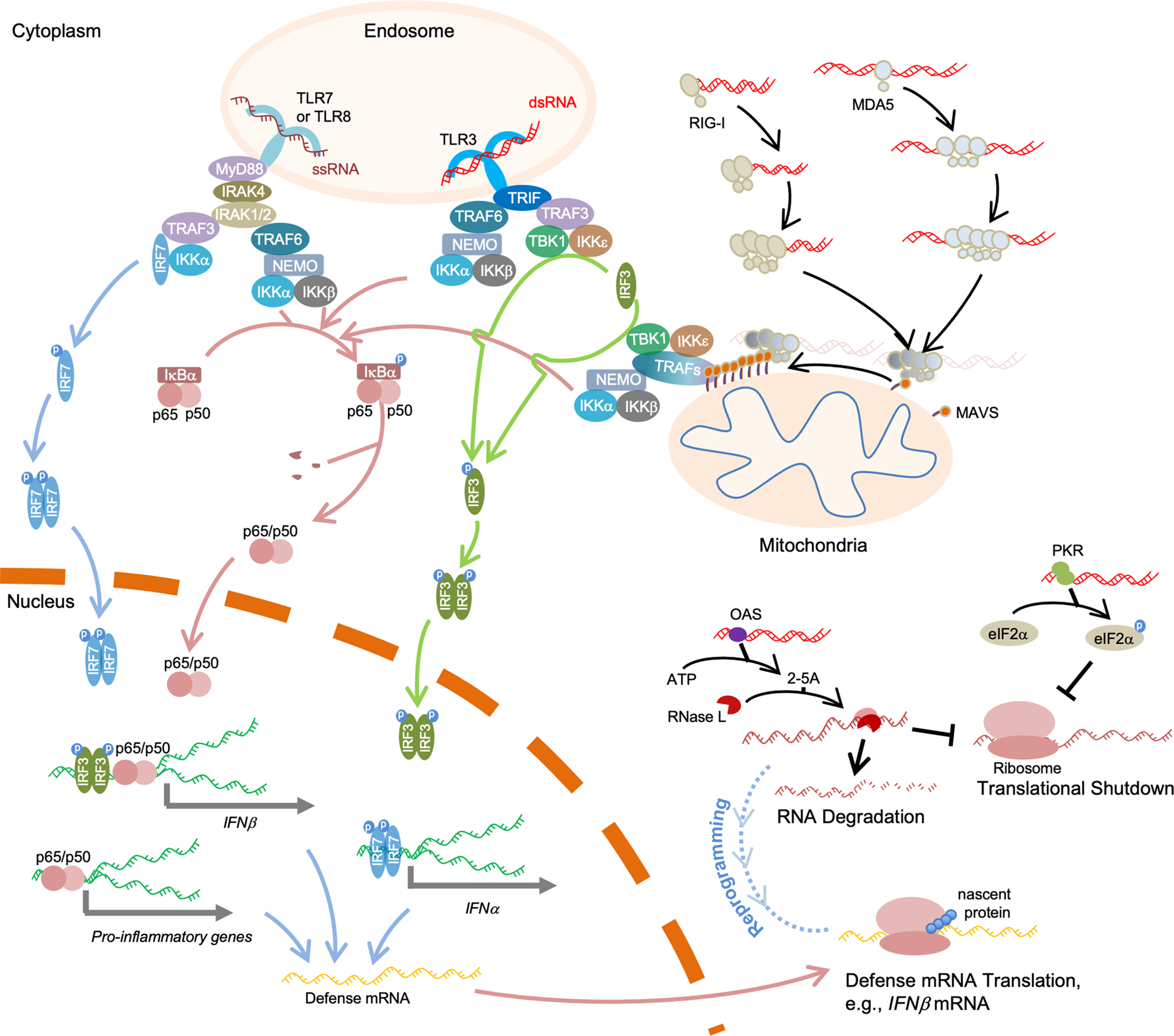
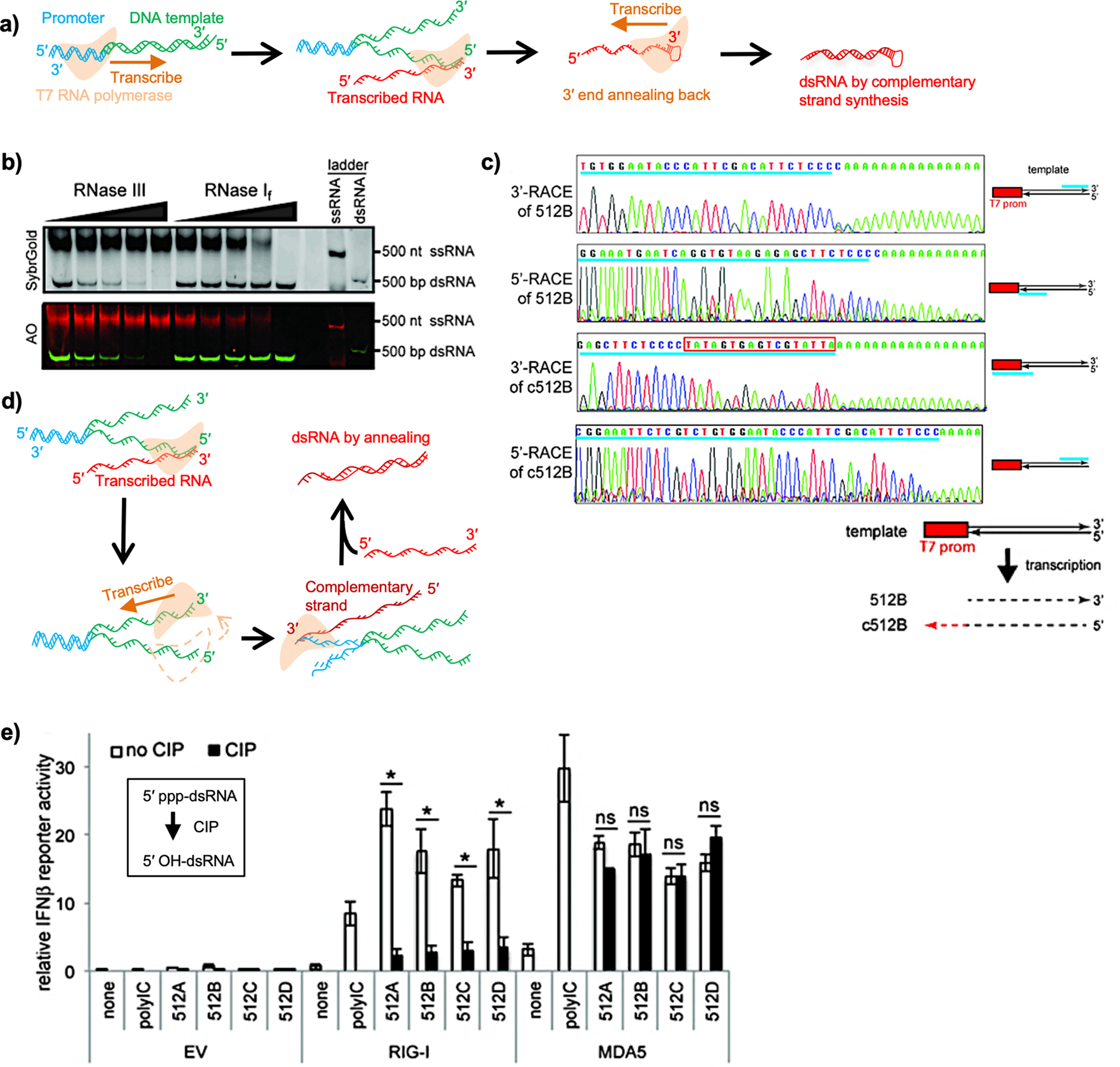
In vitro-transcribed RNAs are emerging as new biologics for therapeutic innovation, as exemplified by their application recently in SARS-CoV-2 vaccinations. RNAs prepared by in vitro transcription (IVT) allow transient expression of proteins of interest, conferring safety over DNA- or virus-mediated gene delivery systems. However, in vitro-transcribed RNAs should be used with caution because of their immunogenicity, which is in part triggered by double-stranded RNA (dsRNA) byproducts during IVT. Cellular innate immune response to dsRNA byproducts can lead to undesirable consequences, including suppression of protein synthesis and cell death, which in turn can detrimentally impact the efficacy of mRNA therapy. Thus, it is critical to understand the nature of IVT byproducts and the mechanisms by which they trigger innate immune responses.Our lab has been investigating the mechanisms by which the innate immune system discriminates between "self" and "nonself" RNA, with the focus on the cytoplasmic dsRNA receptors retinoic acid-inducible gene I (RIG-I) and melanoma differentiation-associated 5 (MDA5). We have biochemically and structurally characterized critical events involving RNA discrimination and signal transduction by RIG-I or MDA5. We have used in vitro-transcribed RNAs as tools to investigate RNA specificity of RIG-I and MDA5, which required optimization of the IVT reaction and purification processes to eliminate the effect of IVT byproducts. In this Account, we summarize our current understanding of RIG-I and MDA5 and IVT reactions and propose future directions for improving IVT as a method to generate both research tools and therapeutics. Other critical proteins in cellular innate immune response to dsRNAs are also discussed. We arrange the contents in the following order: (i) innate immunity sensors for nonself RNA, including the RIG-I-like receptors (RLRs) in the cytosol and the toll-like receptors (TLRs) in the endosome, as well as cytoplasmic dsRNA-responding proteins, including protein kinase R (PKR) and 2',5'-oligoadenylate synthetases (OASes), illustrating the feature of protein-RNA binding and its consequences; (ii) the immunogenicity of IVT byproducts, specifically the generation of dsRNA molecules during IVT; and (iii) methods to reduce IVT RNA immunogenicity, including optimizations of RNA polymerases, reagents, and experimental conditions during IVT and subsequent purification.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
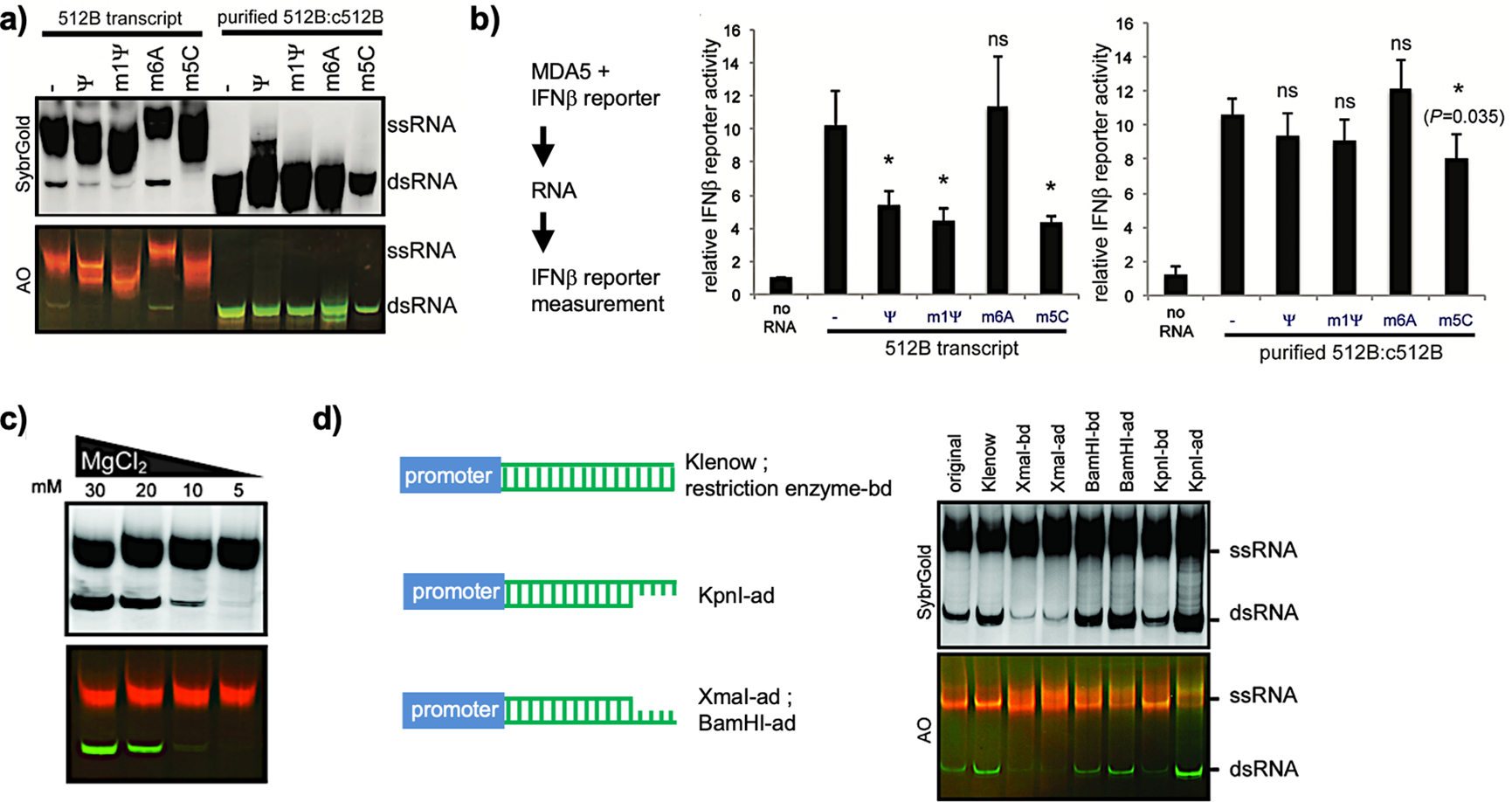
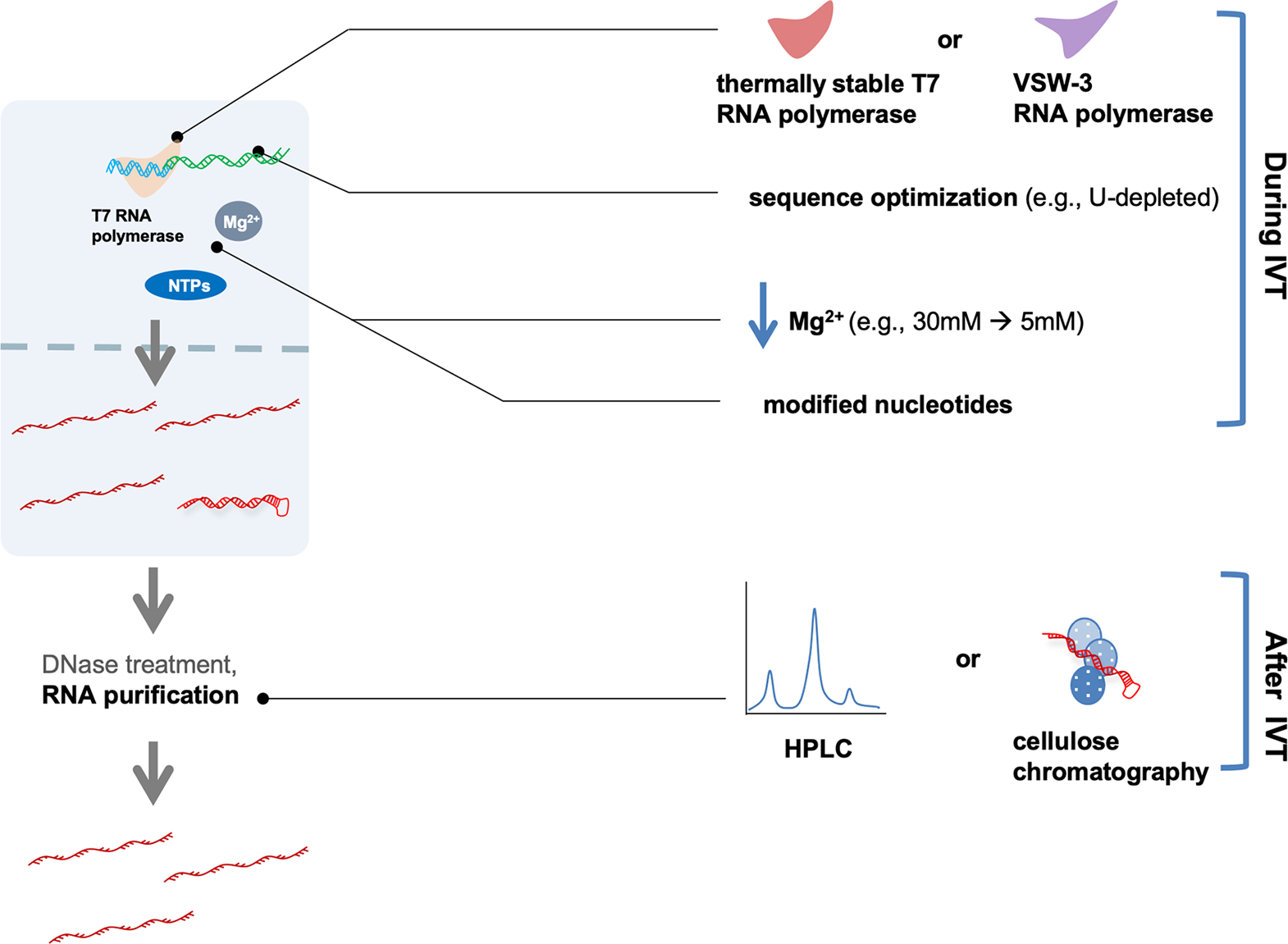
References
-
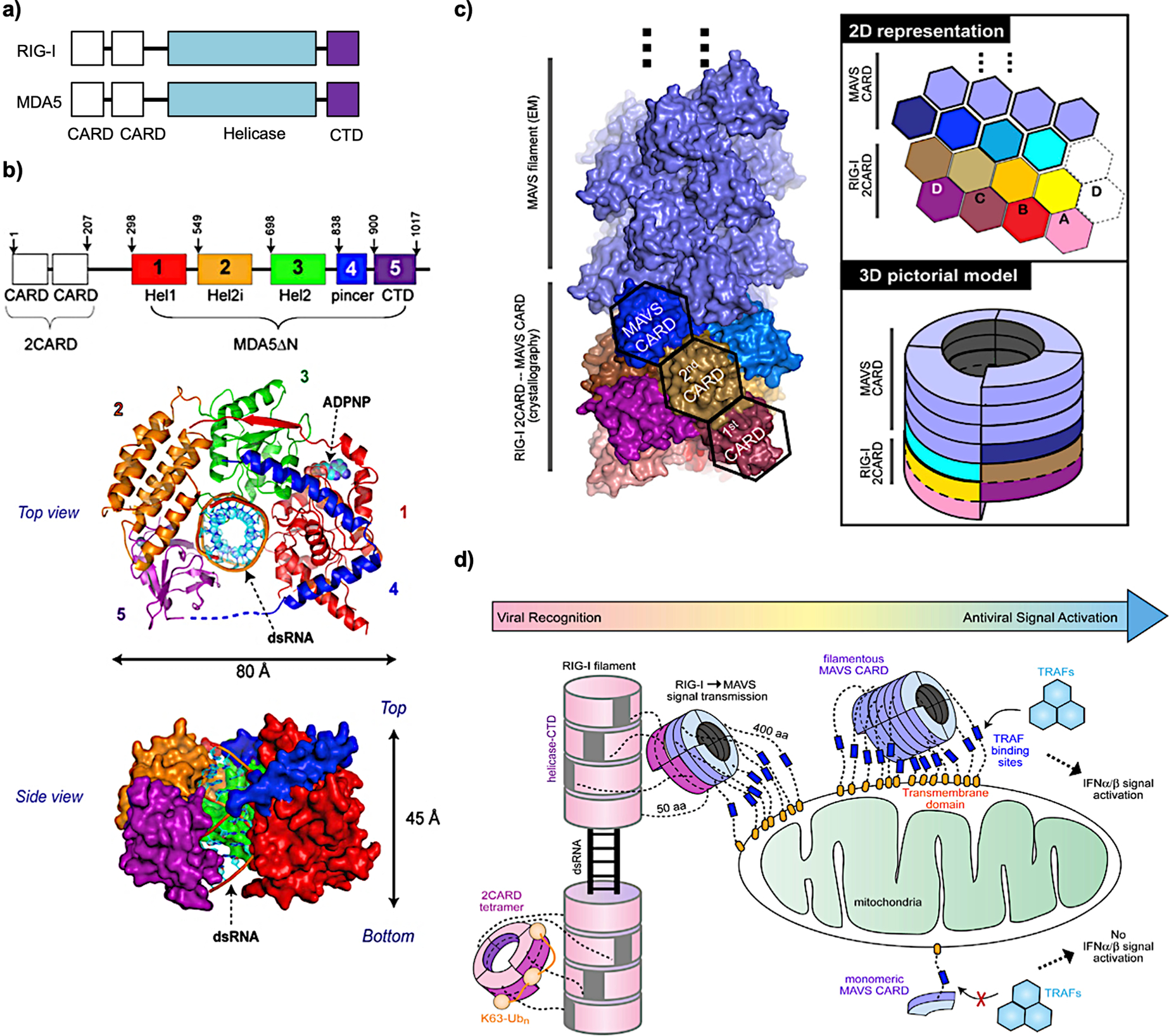
- Wu B; Peisley A; Richards C; Yao H; Zeng X; Lin C; Chu F; Walz T; Hur S Structural Basis for DsRNA Recognition, Filament Formation, and Antiviral Signal Activation by MDA5. Cell 2013, 152, 276–289. - PubMed
-
- Sahin U; Karikó K; Türeci Ö mRNA-Based Therapeutics — Developing a New Class of Drugs. Nat. Rev. Drug Discovery 2014, 13, 759–780. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
