

Simulating Metabolic Flexibility in Low Energy Expenditure Conditions Using Genome-Scale Metabolic Models
- PMID: 34677410
- PMCID: PMC8537358
- DOI: 10.3390/metabo11100695
Simulating Metabolic Flexibility in Low Energy Expenditure Conditions Using Genome-Scale Metabolic Models
Abstract
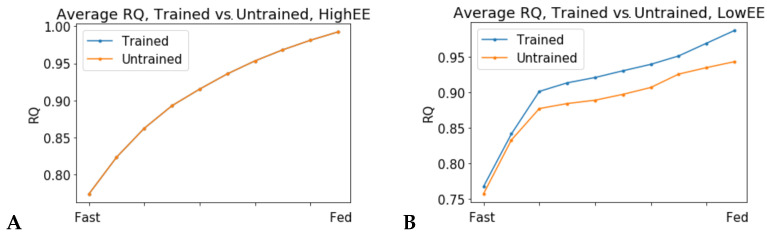
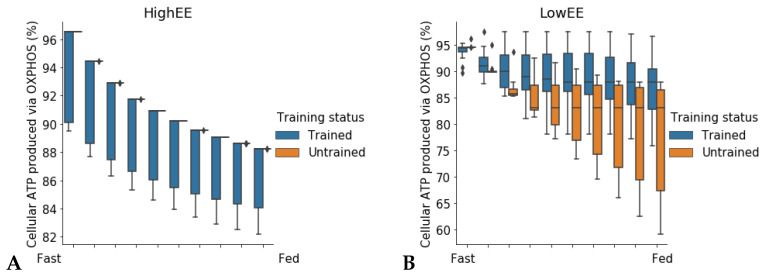
Metabolic flexibility is the ability of an organism to adapt its energy source based on nutrient availability and energy requirements. In humans, this ability has been linked to cardio-metabolic health and healthy aging. Genome-scale metabolic models have been employed to simulate metabolic flexibility by computing the Respiratory Quotient (RQ), which is defined as the ratio of carbon dioxide produced to oxygen consumed, and varies between values of 0.7 for pure fat metabolism and 1.0 for pure carbohydrate metabolism. While the nutritional determinants of metabolic flexibility are known, the role of low energy expenditure and sedentary behavior in the development of metabolic inflexibility is less studied. In this study, we present a new description of metabolic flexibility in genome-scale metabolic models which accounts for energy expenditure, and we study the interactions between physical activity and nutrition in a set of patient-derived models of skeletal muscle metabolism in older adults. The simulations show that fuel choice is sensitive to ATP consumption rate in all models tested. The ability to adapt fuel utilization to energy demands is an intrinsic property of the metabolic network.
Keywords: energy expenditure; metabolic flexibility; respiratory quotient.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




Similar articles
-
Sedentary behaviour is a key determinant of metabolic inflexibility.J Physiol. 2018 Apr 15;596(8):1319-1330. doi: 10.1113/JP273282. Epub 2017 Jul 4. J Physiol. 2018. PMID: 28543022 Free PMC article. Review.
-
Metabolic flexibility and insulin resistance.Am J Physiol Endocrinol Metab. 2008 Nov;295(5):E1009-17. doi: 10.1152/ajpendo.90558.2008. Epub 2008 Sep 2. Am J Physiol Endocrinol Metab. 2008. PMID: 18765680 Free PMC article. Review.
-
Effect of the energy source on changes in energy expenditure, respiratory quotient, and nitrogen balance during total parenteral nutrition in children.Pediatr Res. 1987 Jun;21(6):538-41. doi: 10.1203/00006450-198706000-00006. Pediatr Res. 1987. PMID: 3110723
-
Maternal protein restriction impairs the transcriptional metabolic flexibility of skeletal muscle in adult rat offspring.Br J Nutr. 2014 Aug 14;112(3):328-37. doi: 10.1017/S0007114514000865. Epub 2014 May 14. Br J Nutr. 2014. PMID: 24823946
-
Metabolic flexibility and obesity in children and youth.Obes Rev. 2011 May;12(5):e44-53. doi: 10.1111/j.1467-789X.2010.00812.x. Epub 2010 Oct 26. Obes Rev. 2011. PMID: 20977601 Review.
Cited by
-
Optimizing metabolic health with digital twins.NPJ Aging. 2025 Mar 24;11(1):20. doi: 10.1038/s41514-025-00211-6. NPJ Aging. 2025. PMID: 40128254 Free PMC article. Review.
-
MitoMAMMAL: a genome scale model of mammalian mitochondria predicts cardiac and BAT metabolism.Bioinform Adv. 2024 Nov 5;5(1):vbae172. doi: 10.1093/bioadv/vbae172. eCollection 2025. Bioinform Adv. 2024. PMID: 39758828 Free PMC article.
References
LinkOut - more resources
Full Text Sources
