

The Elusive Cochlear Filter: Wave Origin of Cochlear Cross-Frequency Masking
- PMID: 34677710
- PMCID: PMC8599594
- DOI: 10.1007/s10162-021-00814-2
The Elusive Cochlear Filter: Wave Origin of Cochlear Cross-Frequency Masking
Abstract
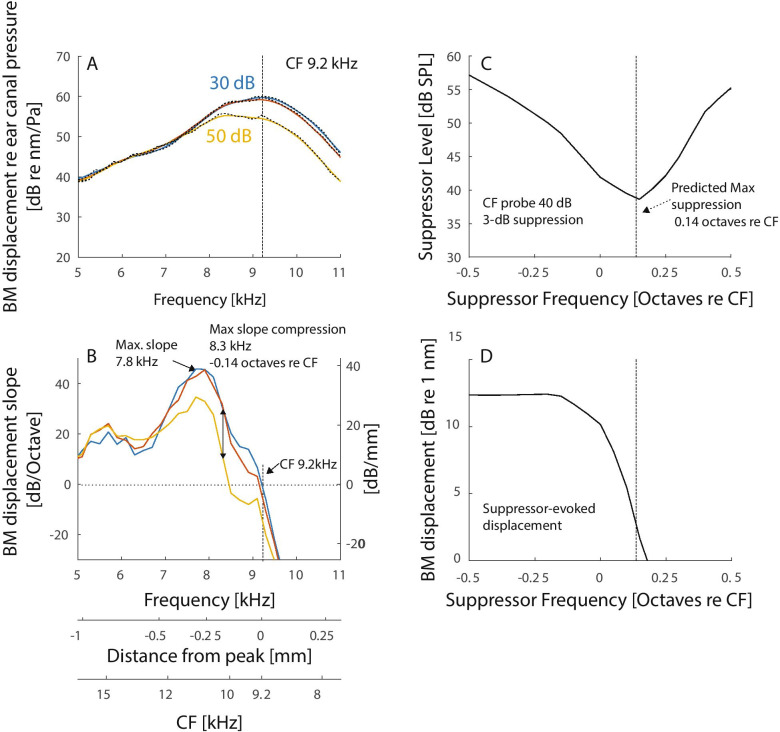
The mammalian cochlea achieves its remarkable sensitivity, frequency selectivity, and dynamic range by spatially segregating the different frequency components of sound via nonlinear processes that remain only partially understood. As a consequence of the wave-based nature of cochlear processing, the different frequency components of complex sounds interact spatially and nonlinearly, mutually suppressing one another as they propagate. Because understanding nonlinear wave interactions and their effects on hearing appears to require mathematically complex or computationally intensive models, theories of hearing that do not deal specifically with cochlear mechanics have often neglected the spatial nature of suppression phenomena. Here we describe a simple framework consisting of a nonlinear traveling-wave model whose spatial response properties can be estimated from basilar-membrane (BM) transfer functions. Without invoking jazzy details of organ-of-Corti mechanics, the model accounts well for the peculiar frequency-dependence of suppression found in two-tone suppression experiments. In particular, our analysis shows that near the peak of the traveling wave, the amplitude of the BM response depends primarily on the nonlinear properties of the traveling wave in more basal (high-frequency) regions. The proposed framework provides perhaps the simplest representation of cochlear signal processing that accounts for the spatially distributed effects of nonlinear wave propagation. Shifting the perspective from local filters to non-local, spatially distributed processes not only elucidates the character of cochlear signal processing, but also has important consequences for interpreting psychophysical experiments.
Keywords: cochlear mechanics; masking; suppression; traveling wave.
© 2021. Association for Research in Otolaryngology.
Figures





Similar articles
-
Amplification and Suppression of Traveling Waves along the Mouse Organ of Corti: Evidence for Spatial Variation in the Longitudinal Coupling of Outer Hair Cell-Generated Forces.J Neurosci. 2019 Mar 6;39(10):1805-1816. doi: 10.1523/JNEUROSCI.2608-18.2019. Epub 2019 Jan 16. J Neurosci. 2019. PMID: 30651330 Free PMC article.
-
The cortilymph wave: Its relation to the traveling wave, auditory-nerve responses, and low-frequency downward glides.Hear Res. 2025 Jun;462:109279. doi: 10.1016/j.heares.2025.109279. Epub 2025 Apr 16. Hear Res. 2025. PMID: 40253777 Review.
-
Age and the human cochlear traveling wave delay.Ear Hear. 1998 Apr;19(2):111-9. doi: 10.1097/00003446-199804000-00003. Ear Hear. 1998. PMID: 9562533
-
Otoacoustic emission estimates of human basilar membrane impulse response duration and cochlear filter tuning.Hear Res. 2016 Dec;342:150-160. doi: 10.1016/j.heares.2016.10.016. Epub 2016 Oct 27. Hear Res. 2016. PMID: 27989947
-
Reflections on the role of a traveling wave along the basilar membrane in view of clinical and experimental findings.Eur Arch Otorhinolaryngol. 2015 Mar;272(3):531-5. doi: 10.1007/s00405-014-3045-z. Epub 2014 Apr 17. Eur Arch Otorhinolaryngol. 2015. PMID: 24740735 Review.
Cited by
-
The spatial buildup of nonlinear compression in the cochlea.Front Cell Neurosci. 2025 Jan 29;18:1450115. doi: 10.3389/fncel.2024.1450115. eCollection 2024. Front Cell Neurosci. 2025. PMID: 39944221 Free PMC article.
-
Overturning the mechanisms of cochlear amplification via area deformations of the organ of Corti.J Acoust Soc Am. 2022 Oct;152(4):2227. doi: 10.1121/10.0014794. J Acoust Soc Am. 2022. PMID: 36319240 Free PMC article.
-
The Shape of Noise to Come: Signal vs. Noise Amplification in the Active Cochlea.AIP Conf Proc. 2024 Feb 27;3062(1):020011. doi: 10.1063/5.0193604. AIP Conf Proc. 2024. PMID: 38516505 Free PMC article.
-
Acoustic estimation of voice roughness.Atten Percept Psychophys. 2025 Jul;87(5):1771-1787. doi: 10.3758/s13414-025-03060-3. Epub 2025 Apr 28. Atten Percept Psychophys. 2025. PMID: 40295423 Free PMC article.
-
Intracochlear overdrive: Characterizing nonlinear wave amplification in the mouse apex.J Acoust Soc Am. 2023 Nov 1;154(5):3414-3428. doi: 10.1121/10.0022446. J Acoust Soc Am. 2023. PMID: 38015028 Free PMC article.
References
-
- Abbas PJ, Sachs MB. Two-tone suppression in auditory-nerve fibers: Extension of a stimulus-response relationship. J Acoust Soc Am. 1976;59(1):112–122. - PubMed
-
- Abdala C, Sininger YS, Ekelid M, Zeng FG. Distortion-product otoacoustic emission suppression tuning curves in human adults and neonates. Hear Res. 1996;98:38–53. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
