

The two-Cys-type TetR repressor GbaA confers resistance under disulfide and electrophile stress in Staphylococcus aureus
- PMID: 34678418
- PMCID: PMC8693949
- DOI: 10.1016/j.freeradbiomed.2021.10.024
The two-Cys-type TetR repressor GbaA confers resistance under disulfide and electrophile stress in Staphylococcus aureus
Abstract
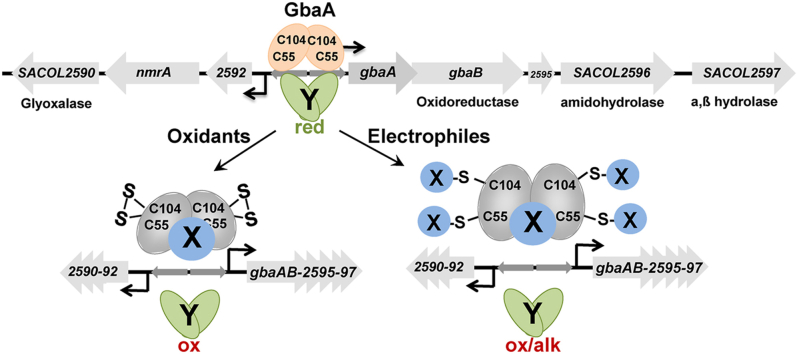
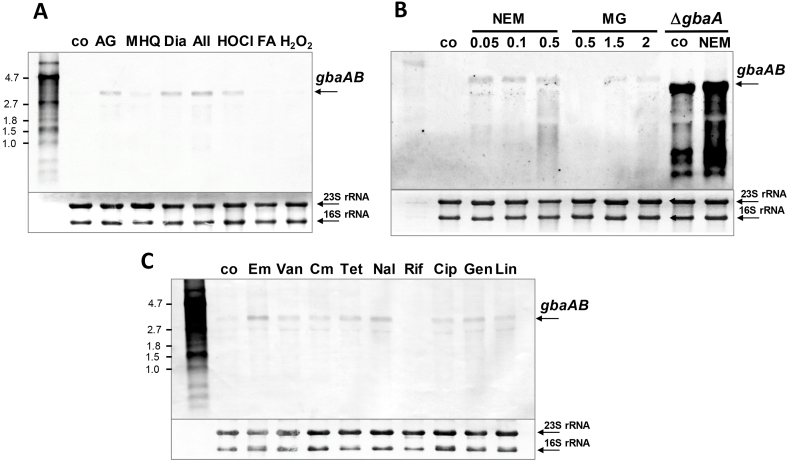
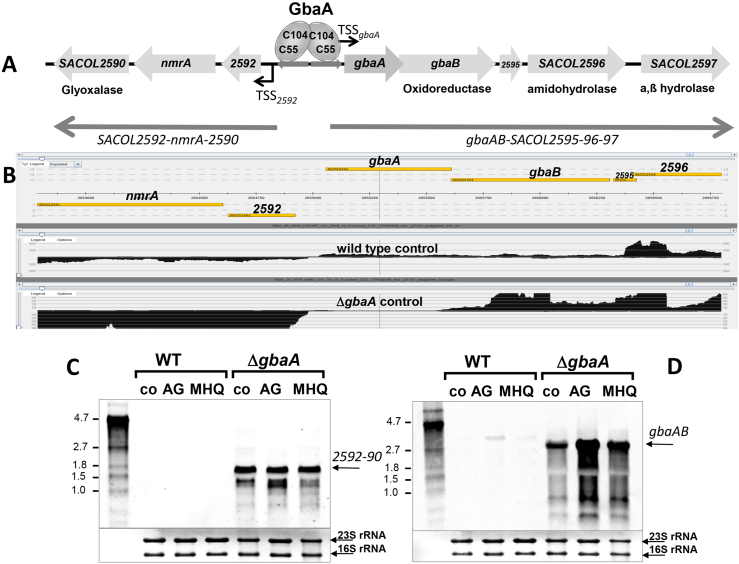
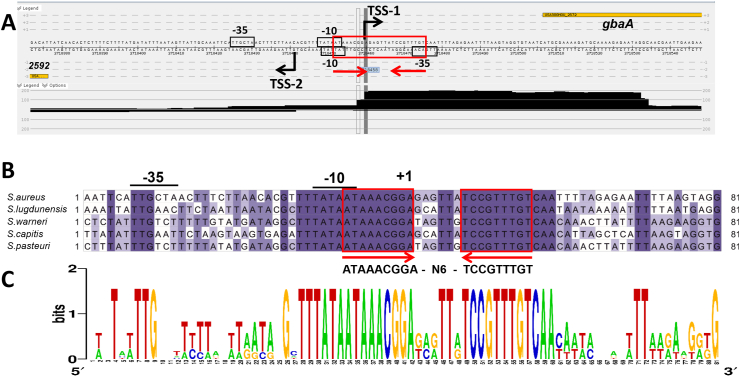
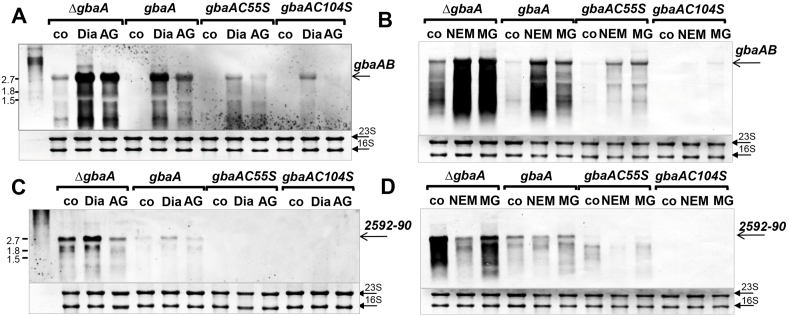
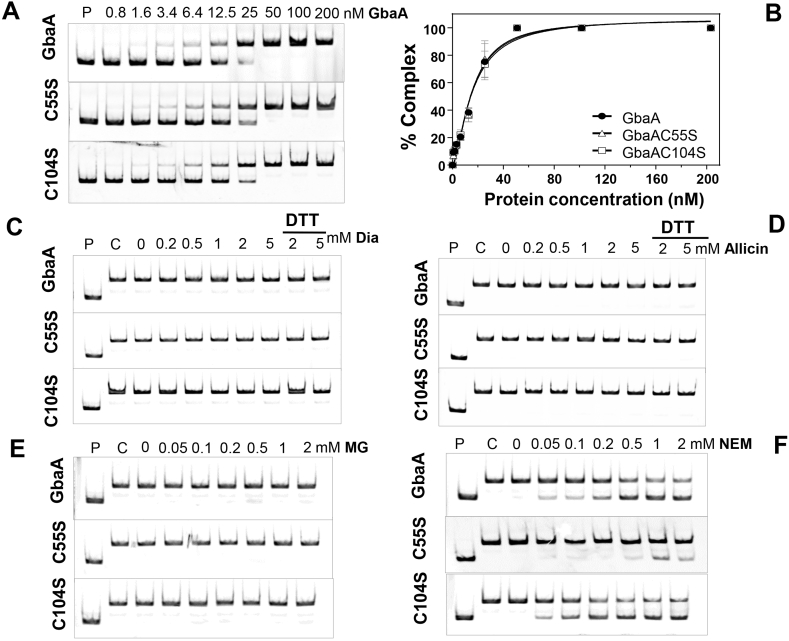
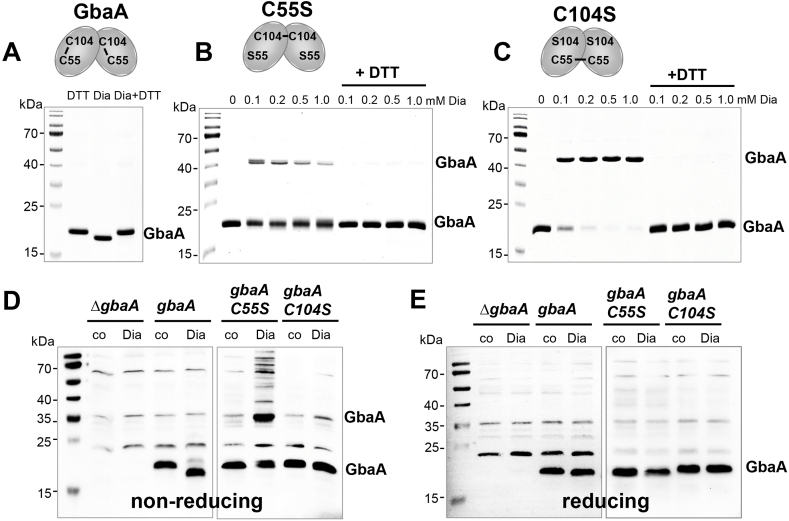
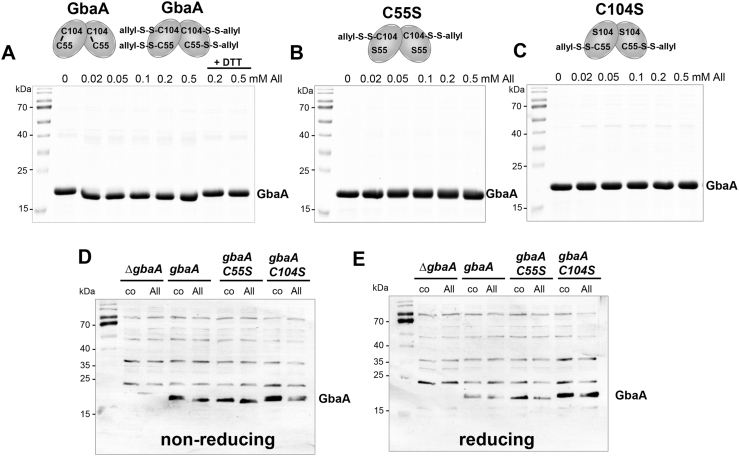
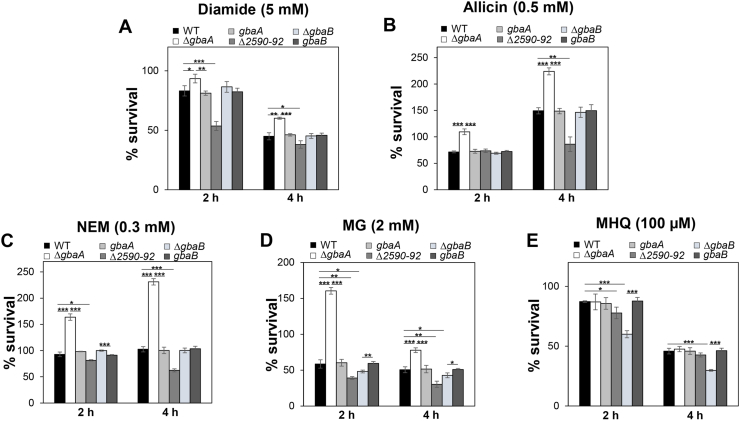
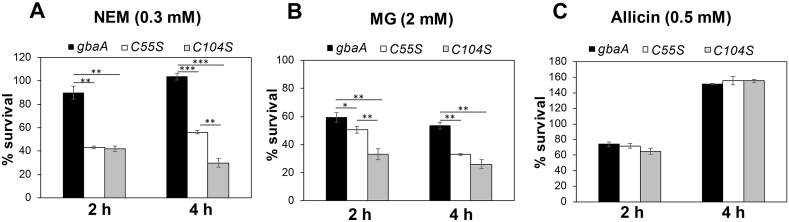
Staphylococcus aureus has to cope with oxidative and electrophile stress during host-pathogen interactions. The TetR-family repressor GbaA was shown to sense electrophiles, such as N-ethylmaleimide (NEM) via monothiol mechanisms of the two conserved Cys55 or Cys104 residues in vitro. In this study, we further investigated the regulation and function of the GbaA repressor and its Cys residues in S. aureus COL. The GbaA-controlled gbaAB-SACOL2595-97 and SACOL2592-nmrA-2590 operons were shown to respond only weakly 3-10-fold to oxidants, electrophiles or antibiotics in S. aureus COL, but are 57-734-fold derepressed in the gbaA deletion mutant, indicating that the physiological inducer is still unknown. Moreover, the gbaA mutant remained responsive to disulfide and electrophile stress, pointing to additional redox control mechanisms of both operons. Thiol-stress induction of the GbaA regulon was strongly diminished in both single Cys mutants, supporting that both Cys residues are required for redox-sensing in vivo. While GbaA and the single Cys mutants are reversible oxidized under diamide and allicin stress, these thiol switches did not affect the DNA binding activity. The repressor activity of GbaA could be only partially inhibited with NEM in vitro. Survival assays revealed that the gbaA mutant confers resistance under diamide, allicin, NEM and methylglyoxal stress, which was mediated by the SACOL2592-90 operon encoding for a putative glyoxalase and oxidoreductase. Altogether, our results support that the GbaA repressor functions in the defense against oxidative and electrophile stress in S. aureus. GbaA represents a 2-Cys-type redox sensor, which requires another redox-sensing regulator and an unknown thiol-reactive ligand for full derepression of the GbaA regulon genes.
Keywords: Allicin; Diamide; Electrophiles; GbaA; Staphylococcus aureus; Thiol switches.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare that there is no conflict of interest.
Figures
Similar articles
-
Staphylococcus aureus Glucose-Induced Biofilm Accessory Protein A (GbaA) Is a Monothiol-Dependent Electrophile Sensor.Biochemistry. 2020 Aug 11;59(31):2882-2895. doi: 10.1021/acs.biochem.0c00347. Epub 2020 Jul 29. Biochemistry. 2020. PMID: 32697076 Free PMC article.
-
The MarR/DUF24-Family QsrR Repressor Senses Quinones and Oxidants by Thiol Switch Mechanisms in Staphylococcus aureus.Antioxid Redox Signal. 2023 May;38(13-15):877-895. doi: 10.1089/ars.2022.0090. Epub 2022 Nov 28. Antioxid Redox Signal. 2023. PMID: 36242097
-
Redox-Sensing Under Hypochlorite Stress and Infection Conditions by the Rrf2-Family Repressor HypR in Staphylococcus aureus.Antioxid Redox Signal. 2018 Sep 1;29(7):615-636. doi: 10.1089/ars.2017.7354. Epub 2018 Jan 30. Antioxid Redox Signal. 2018. PMID: 29237286 Free PMC article.
-
Thiol-based redox switches in the major pathogen Staphylococcus aureus.Biol Chem. 2020 Nov 23;402(3):333-361. doi: 10.1515/hsz-2020-0272. Print 2021 Feb 23. Biol Chem. 2020. PMID: 33544504 Review.
-
Thiol-based redox switches in prokaryotes.Biol Chem. 2015 May;396(5):415-44. doi: 10.1515/hsz-2015-0102. Biol Chem. 2015. PMID: 25720121 Free PMC article. Review.
Cited by
-
In vitro activities of licochalcone A against planktonic cells and biofilm of Enterococcus faecalis.Front Microbiol. 2022 Oct 21;13:970901. doi: 10.3389/fmicb.2022.970901. eCollection 2022. Front Microbiol. 2022. PMID: 36338074 Free PMC article.
-
Transcription-replication interactions reveal bacterial genome regulation.Nature. 2024 Feb;626(7999):661-669. doi: 10.1038/s41586-023-06974-w. Epub 2024 Jan 24. Nature. 2024. PMID: 38267581 Free PMC article.
-
Transcription-replication interactions reveal principles of bacterial genome regulation.Res Sq [Preprint]. 2023 Mar 31:rs.3.rs-2724389. doi: 10.21203/rs.3.rs-2724389/v1. Res Sq. 2023. Update in: Nature. 2024 Feb;626(7999):661-669. doi: 10.1038/s41586-023-06974-w. PMID: 37034646 Free PMC article. Updated. Preprint.
-
How to Recruit a Promiscuous Enzyme to Serve a New Function.Biochemistry. 2023 Jan 17;62(2):300-308. doi: 10.1021/acs.biochem.2c00249. Epub 2022 Jun 21. Biochemistry. 2023. PMID: 35729117 Free PMC article.
-
Bacterial Metallostasis: Metal Sensing, Metalloproteome Remodeling, and Metal Trafficking.Chem Rev. 2024 Dec 25;124(24):13574-13659. doi: 10.1021/acs.chemrev.4c00264. Epub 2024 Dec 10. Chem Rev. 2024. PMID: 39658019 Free PMC article. Review.
References
-
- Lowy F.D. Staphylococcus aureus infections. N. Engl. J. Med. 1998;339(8):520–532. - PubMed
-
- Boucher H.W., Corey G.R. Epidemiology of methicillin-resistant Staphylococcus aureus. Clin. Infect. Dis. 2008;46(Suppl 5):S344–S349. - PubMed
-
- Archer G.L. Staphylococcus aureus: a well-armed pathogen. Clin. Infect. Dis. 1998;26(5):1179–1181. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
