

The Peculiar Estrogenicity of Diethyl Phthalate: Modulation of Estrogen Receptor α Activities in the Proliferation of Breast Cancer Cells
- PMID: 34678933
- PMCID: PMC8538674
- DOI: 10.3390/toxics9100237
The Peculiar Estrogenicity of Diethyl Phthalate: Modulation of Estrogen Receptor α Activities in the Proliferation of Breast Cancer Cells
Abstract
Phthalates comprise a group of synthetic chemicals present in the environment because of their wide use as plasticizers and as additives in products for personal care. Among others, diethyl phthalate (DEP) is largely used in products for infants, children, and adults, in which its exposure has been correlated with an increased risk of breast cancer. The adverse health outcomes deriving from phthalate exposure have been associated with their activity as endocrine disruptors (EDCs) of the steroid and thyroid hormone signaling by affecting developmental and reproductive health, and even carcinogenicity. However, the estrogen disruptor activities of DEP are still controversial, and the mechanism at the root of the estrogenic-disrupting action of DEP remains to be clarified. Here, we evaluated the DEP mechanism of action on the activation status of estrogen receptor α (ERα) by analyzing the receptor's phosphorylation as well as both nuclear and extra-nuclear pathways triggered by the receptor to modulate the proliferation of breast cancer cells. Although DEP does not bind to ERα, our results suggest that this phthalate ester exerts multiple parallel interactions with ERα signaling and emphasize the importance to determine an appropriate battery of in vitro methods that will include specific molecular mechanisms involved in the endocrine disruption.
Keywords: breast cancer cell proliferation; diethyl phthalate; endocrine disruptors; estrogen receptor α; signal transduction pathways.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
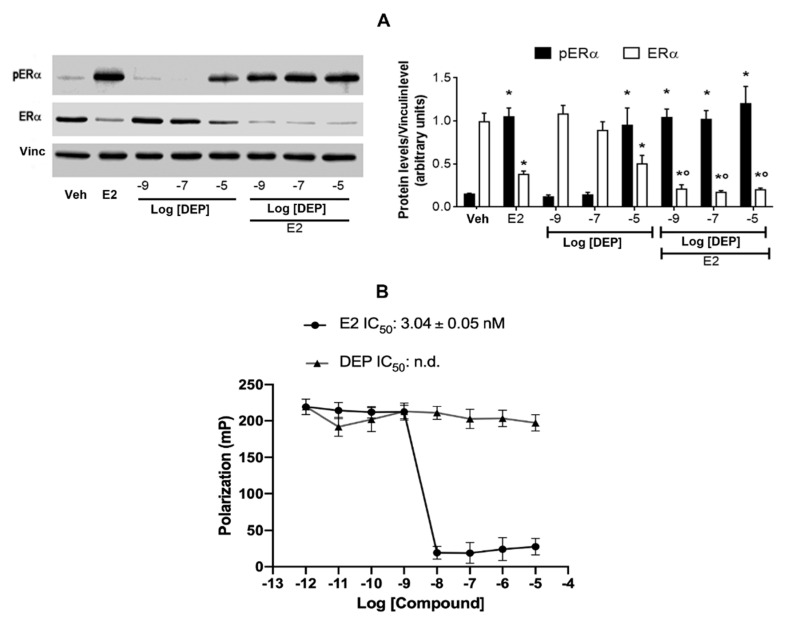
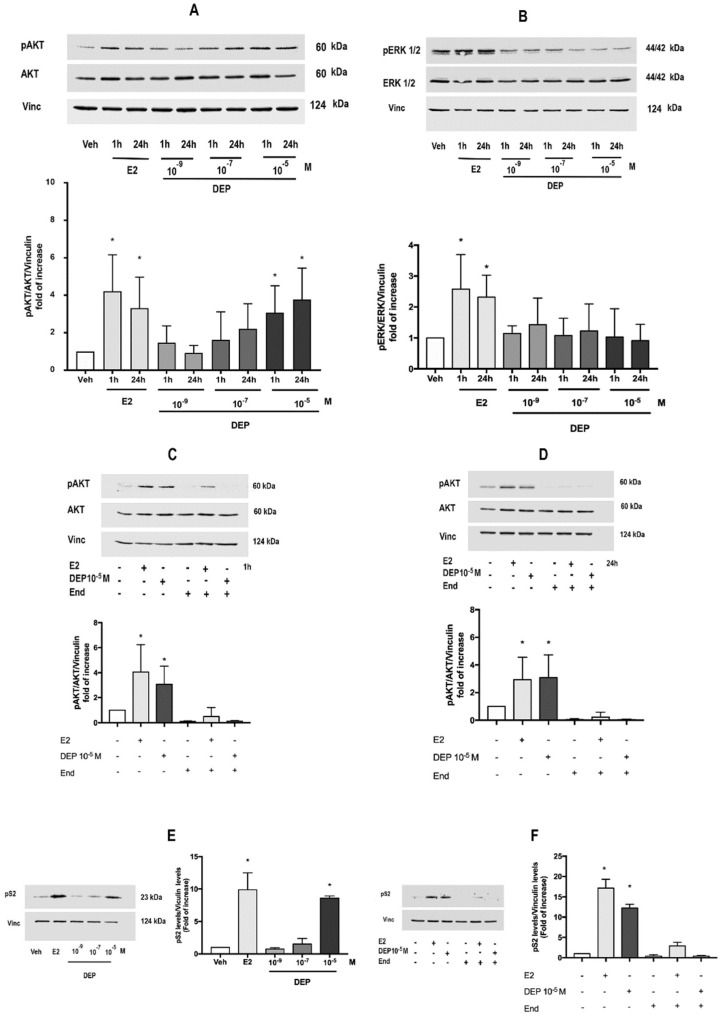
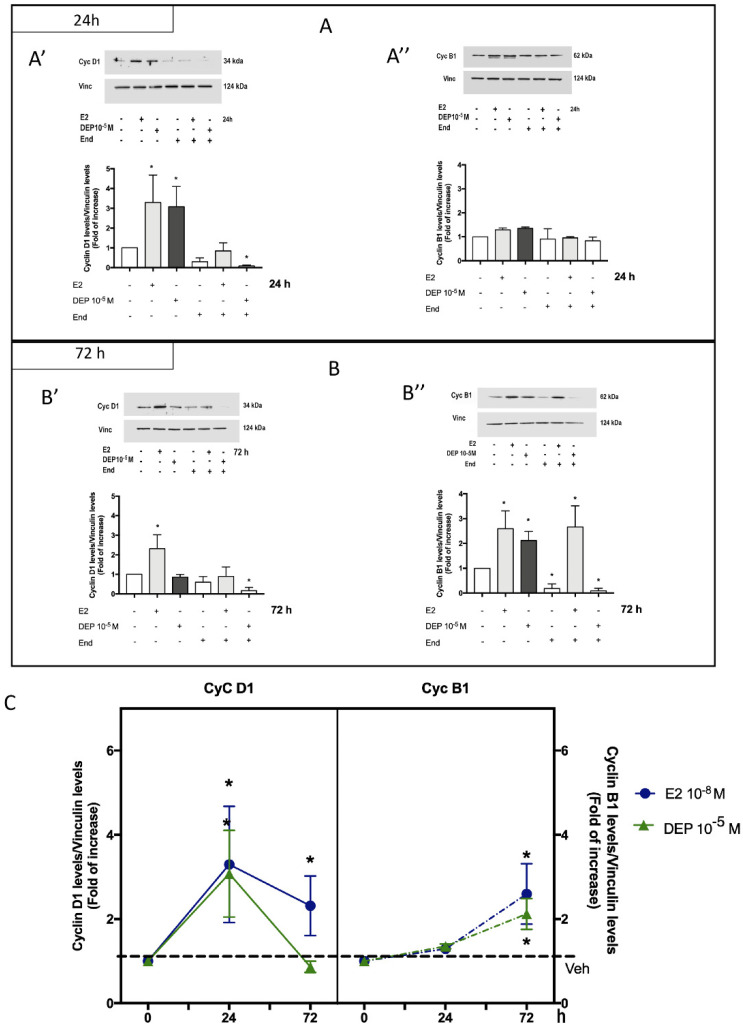
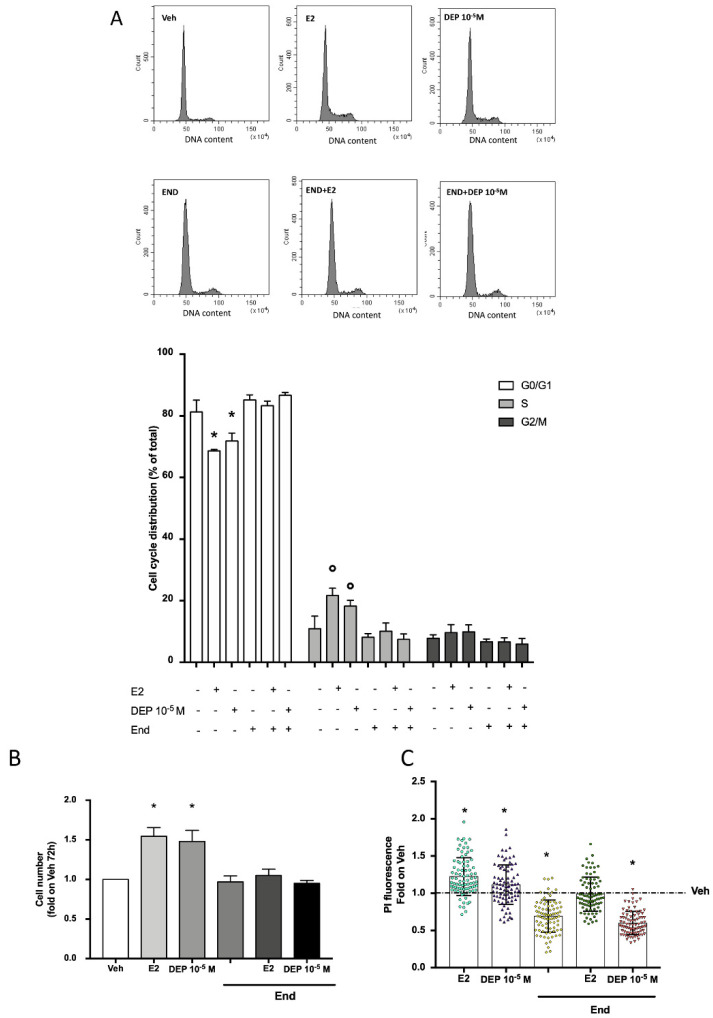
References
Grants and funding
LinkOut - more resources
Full Text Sources
