

Sterols, Oxysterols, and Accessible Cholesterol: Signalling for Homeostasis, in Immunity and During Development
- PMID: 34690800
- PMCID: PMC8531217
- DOI: 10.3389/fphys.2021.723224
Sterols, Oxysterols, and Accessible Cholesterol: Signalling for Homeostasis, in Immunity and During Development
Abstract
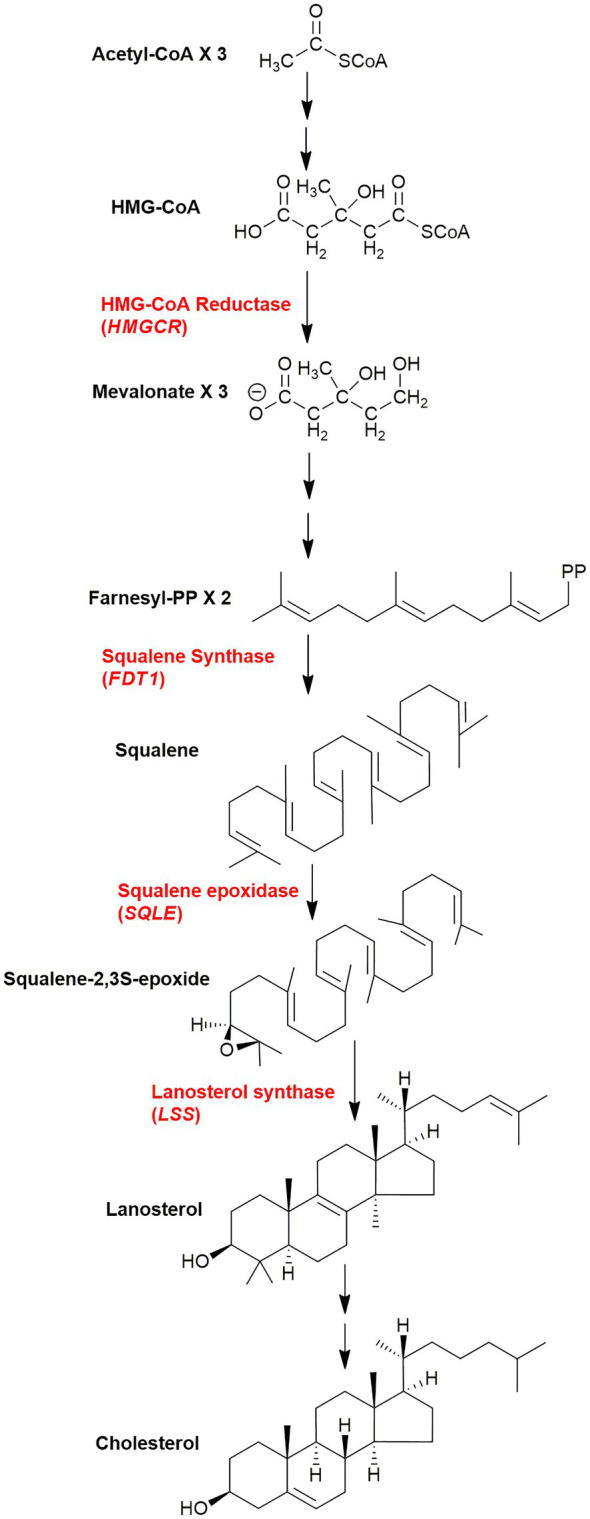
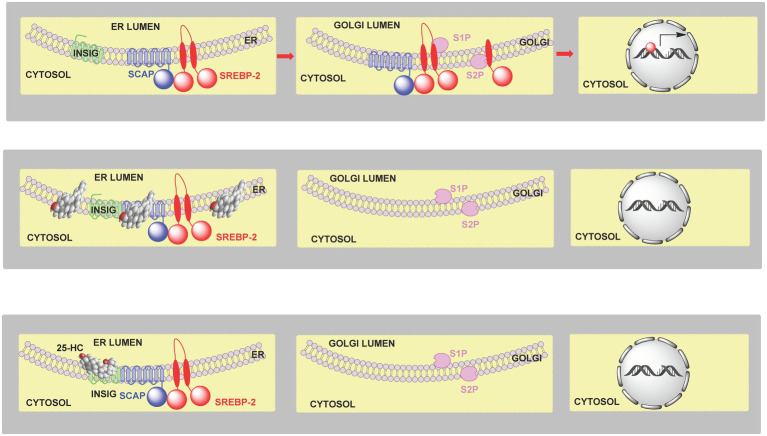
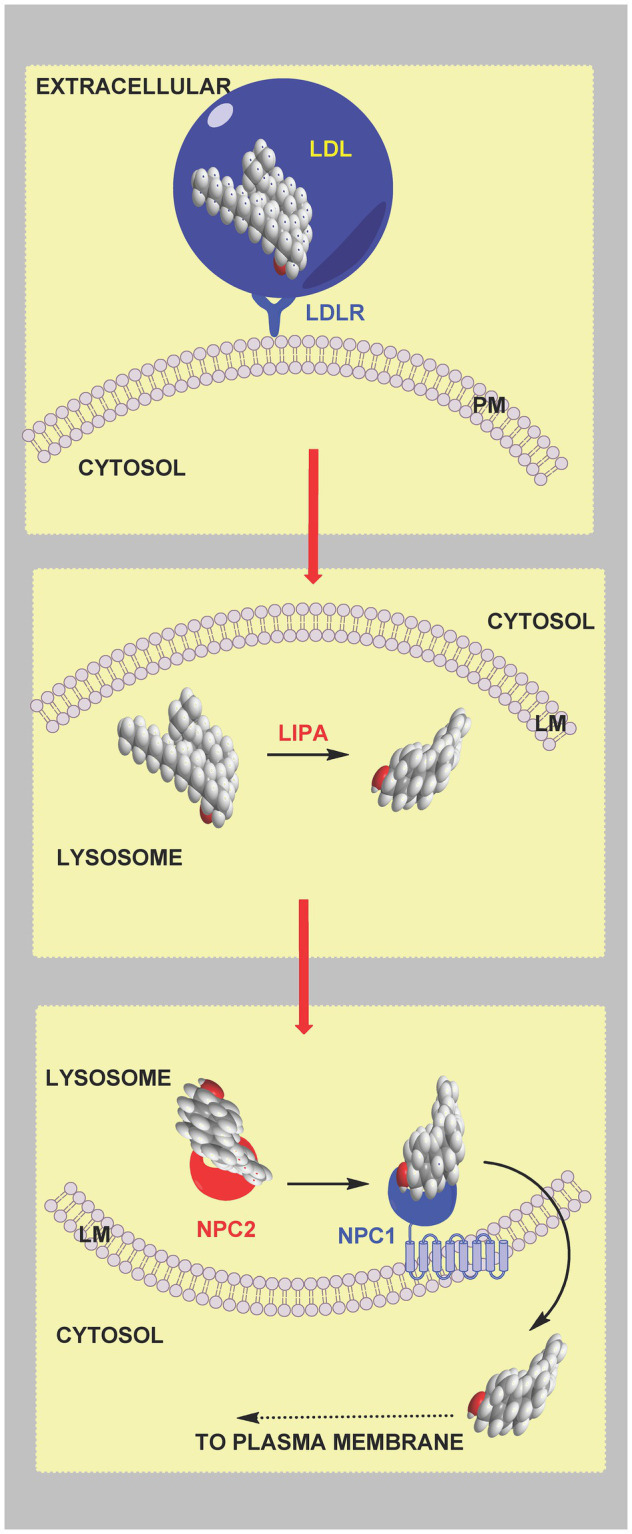
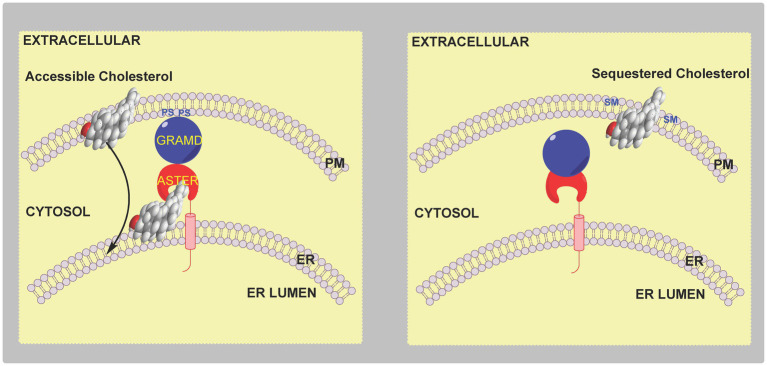
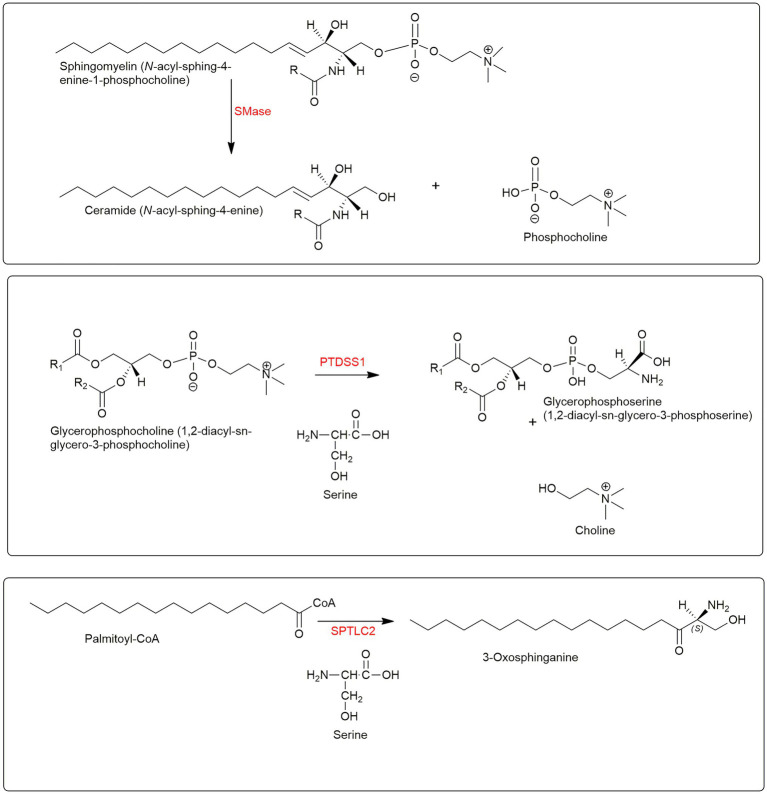
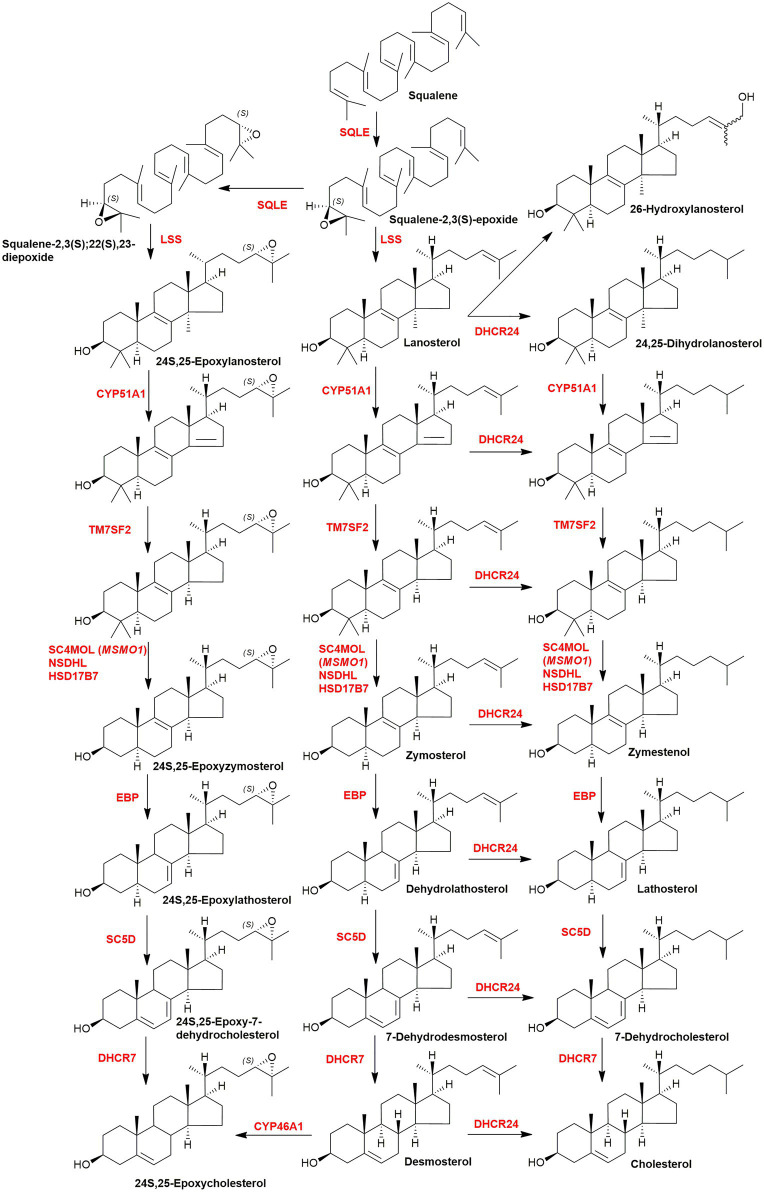
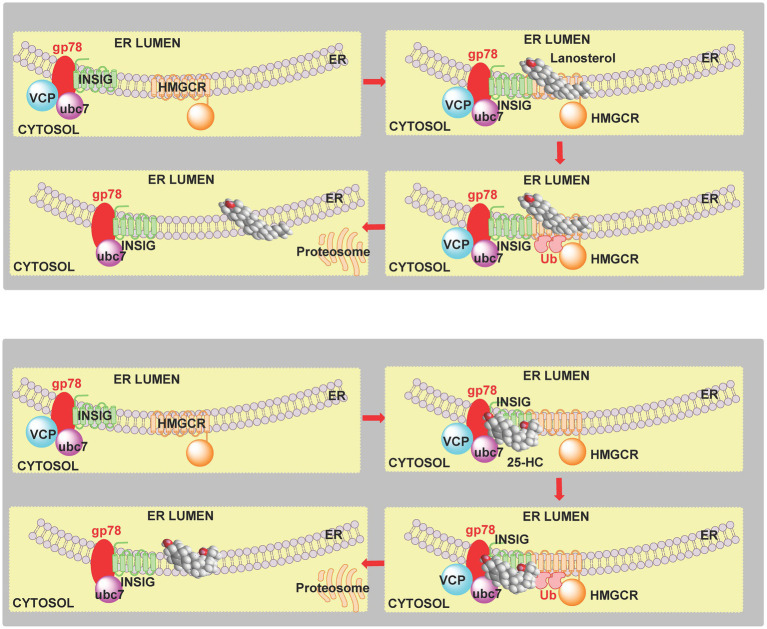
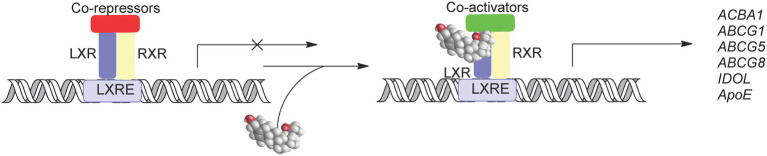
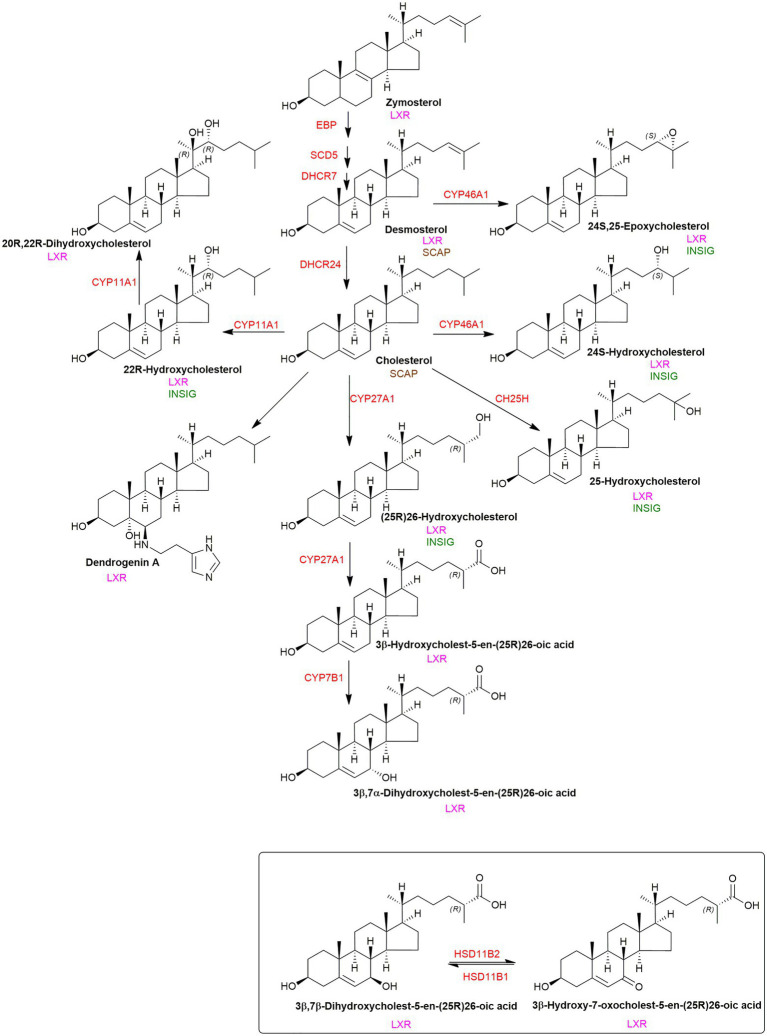
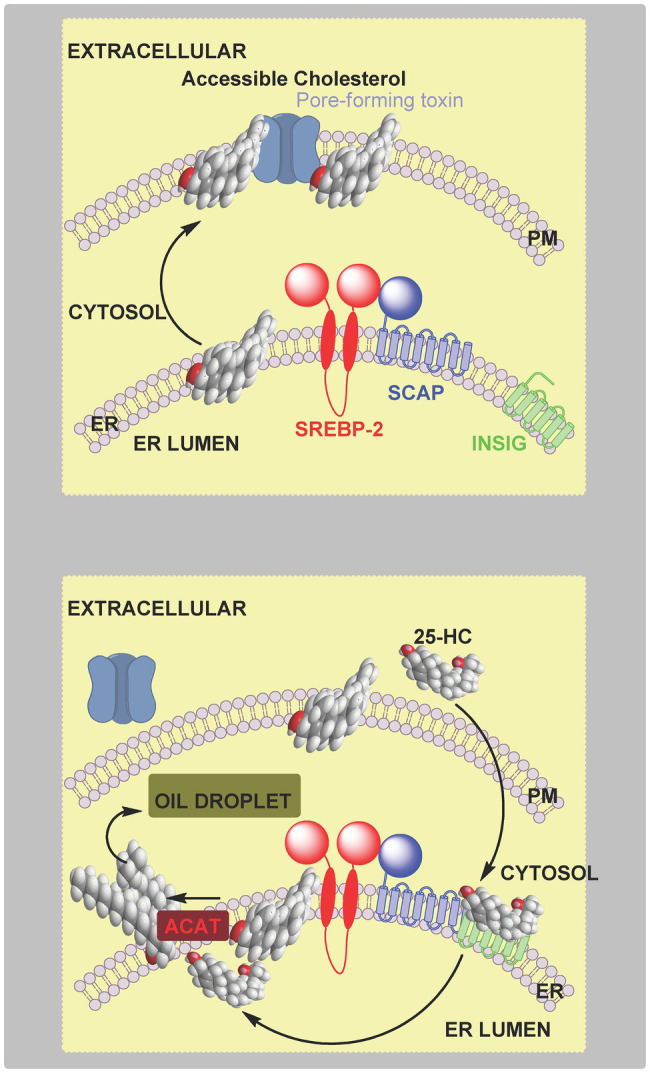
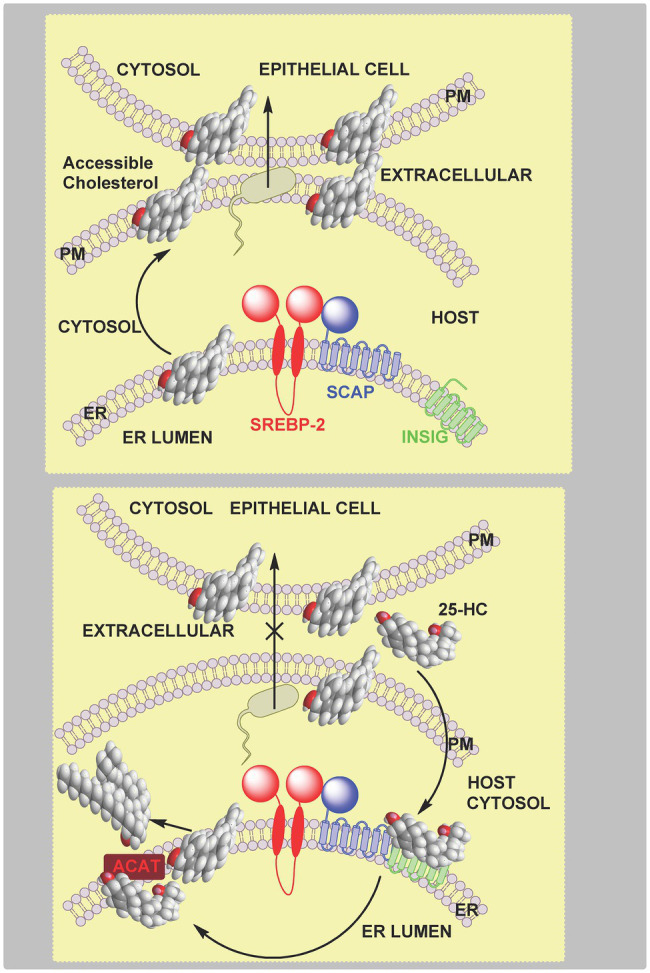
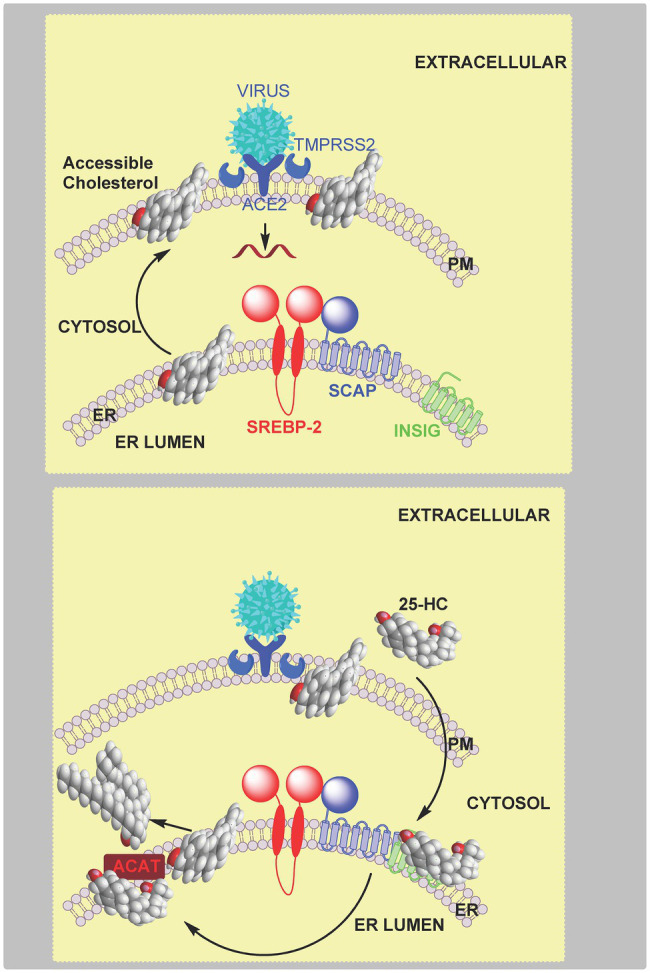
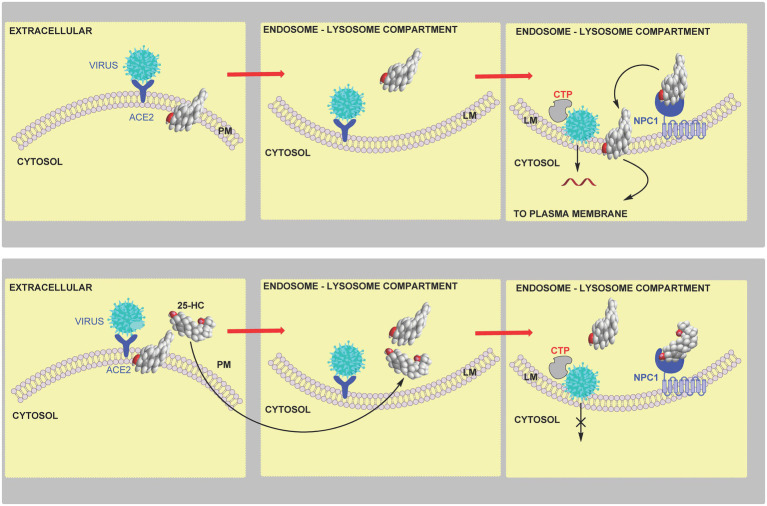
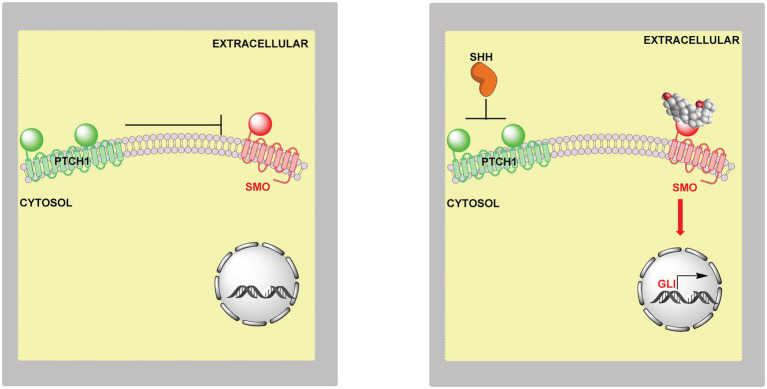
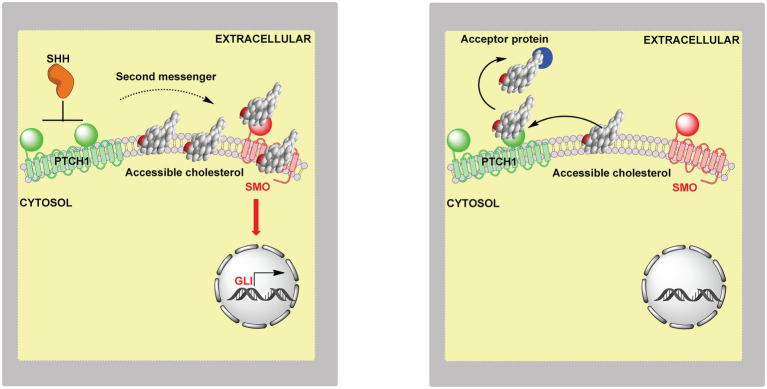
In this article we discuss the concept of accessible plasma membrane cholesterol and its involvement as a signalling molecule. Changes in plasma membrane accessible cholesterol, although only being minor in the context of total cholesterol plasma membrane cholesterol and total cell cholesterol, are a key regulator of overall cellular cholesterol homeostasis by the SREBP pathway. Accessible cholesterol also provides the second messenger between patched 1 and smoothened in the hedgehog signalling pathway important during development, and its depletion may provide a mechanism of resistance to microbial pathogens including SARS-CoV-2. We revise the hypothesis that oxysterols are a signalling form of cholesterol, in this instance as a rapidly acting and paracrine version of accessible cholesterol.
Keywords: 25-hydroxycholesterol; HMG-CoA reductase; INSIG; SARS-CoV-2; SREBP pathway; cholesterol dependent cytolysin; hedgehog signalling pathway.
Copyright © 2021 Griffiths and Wang.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
-
- Abdel-Khalik J., Hearn T., Dickson A. L., Crick P. J., Yutuc E., Austin-Muttitt K., et al. (2020). Bile acid biosynthesis in Smith-Lemli-Opitz syndrome bypassing cholesterol: potential importance of pathway intermediates. J. Steroid Biochem. Mol. Biol. 206:105794. doi: 10.1016/j.jsbmb.2020.105794, PMID: - DOI - PMC - PubMed
-
- Babiker A., Andersson O., Lindblom D., van der Linden J., Wiklund B., Lutjohann D., et al. (1999). Elimination of cholesterol as cholestenoic acid in human lung by sterol 27-hydroxylase: evidence that most of this steroid in the circulation is of pulmonary origin. J. Lipid Res. 40, 1417–1425. doi: 10.1016/S0022-2275(20)33383-6, PMID: - DOI - PubMed
-
- Bauman D. R., Bitmansour A. D., McDonald J. G., Thompson B. M., Liang G., Russell D. W. (2009). 25-Hydroxycholesterol secreted by macrophages in response to toll-like receptor activation suppresses immunoglobulin A production. Proc. Natl. Acad. Sci. U. S. A. 106, 16764–16769. doi: 10.1073/pnas.0909142106, PMID: - DOI - PMC - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Miscellaneous