

Super-enhancer-associated TMEM44-AS1 aggravated glioma progression by forming a positive feedback loop with Myc
- PMID: 34696771
- PMCID: PMC8543865
- DOI: 10.1186/s13046-021-02129-9
Super-enhancer-associated TMEM44-AS1 aggravated glioma progression by forming a positive feedback loop with Myc
Abstract
Background: Long non-coding RNAs (lncRNAs) have been considered as one type of gene expression regulator for cancer development, but it is not clear how these are regulated. This study aimed to identify a specific lncRNA that promotes glioma progression.
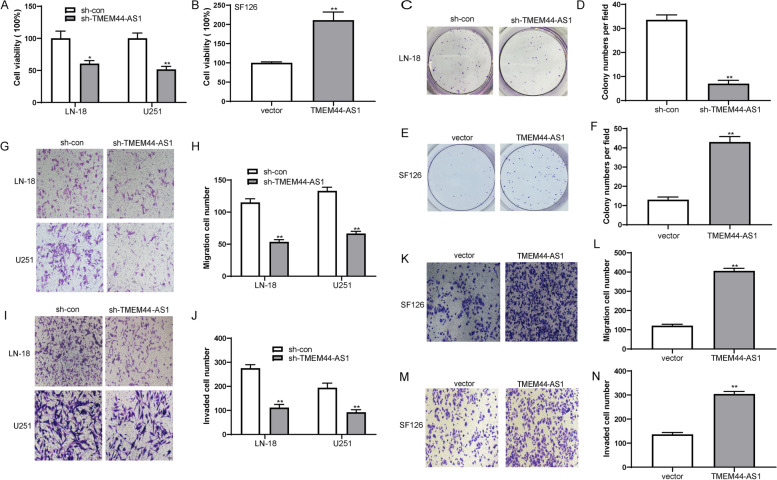
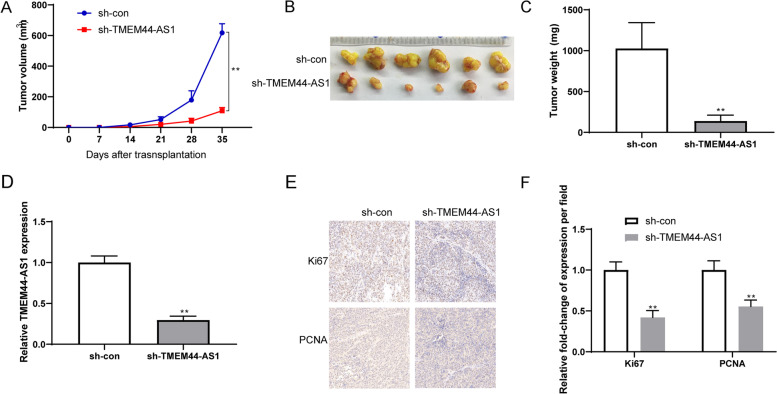
Methods: RNA sequencing (RNA-seq) and quantitative real-time PCR were performed to screen differentially expressed genes. CCK-8, transwell migration, invasion assays, and a mouse xenograft model were performed to determine the functions of TMEM44-AS1. Co-IP, ChIP, Dual-luciferase reporter assays, RNA pulldown, and RNA immunoprecipitation assays were performed to study the molecular mechanism of TMEM44-AS1 and the downstream target.
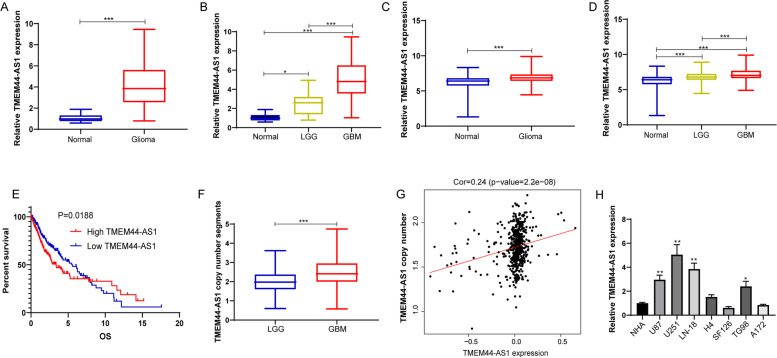
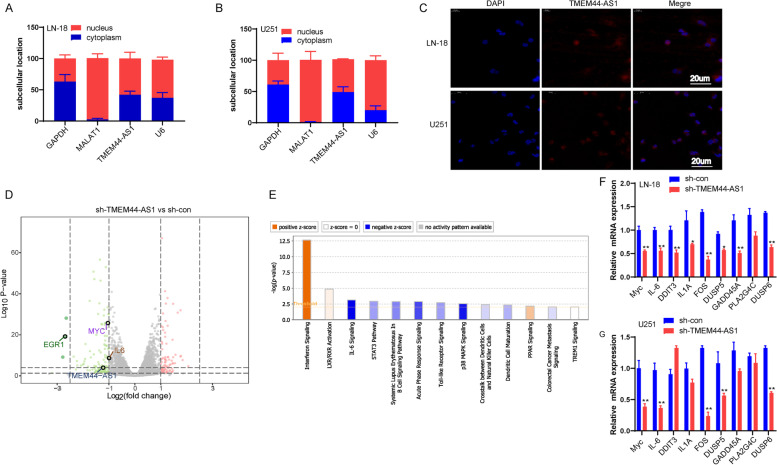
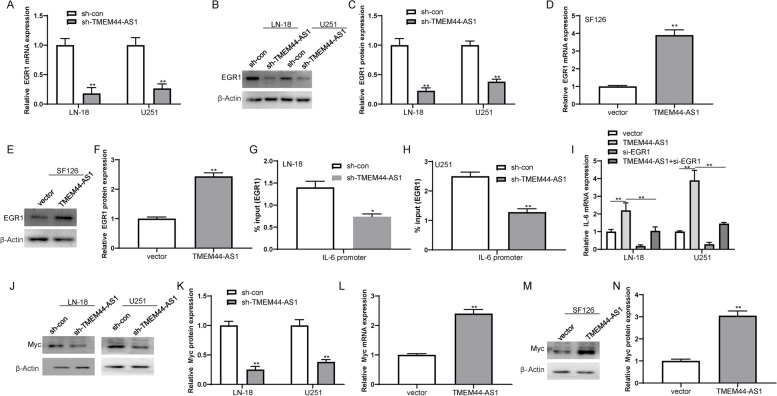
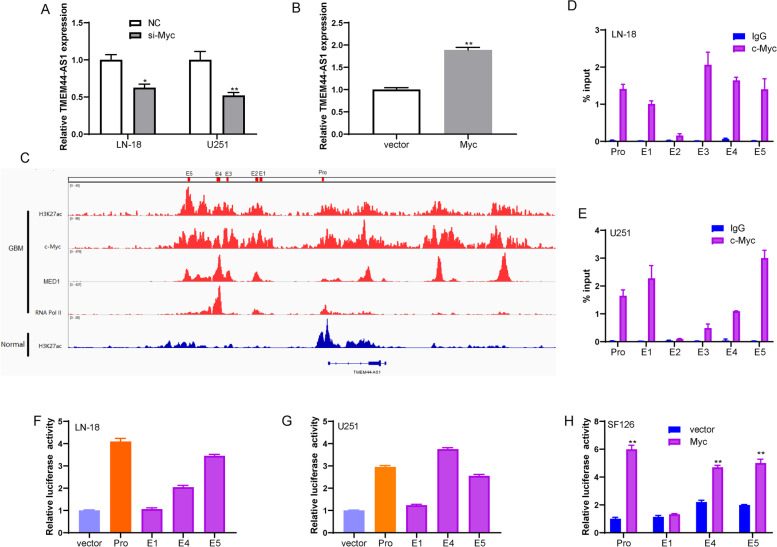
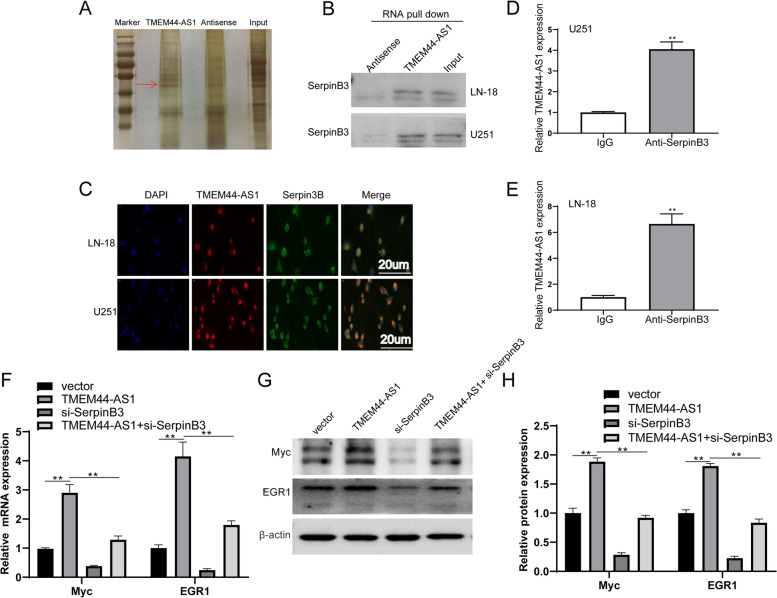
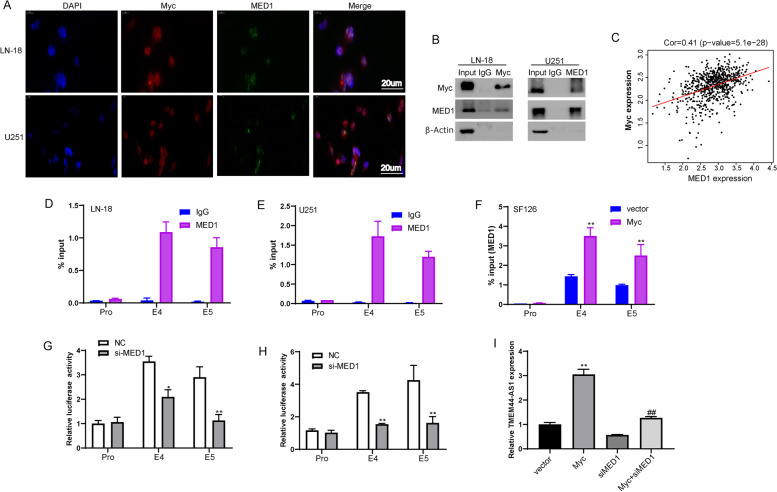
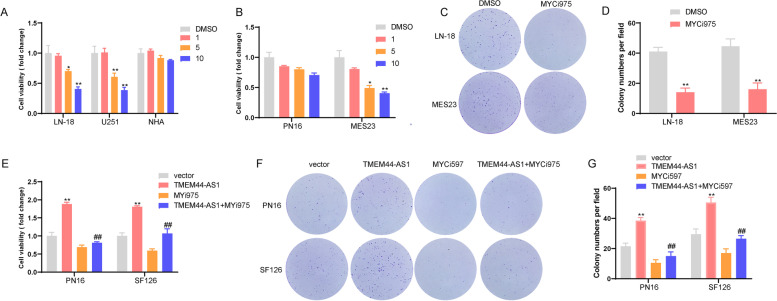
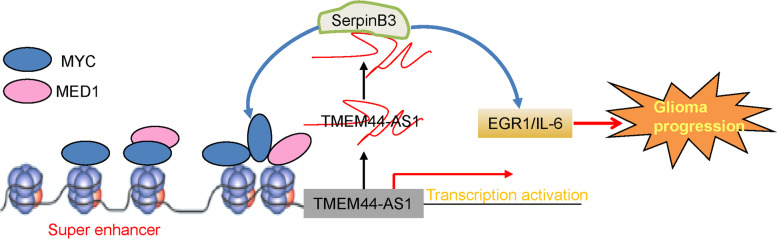
Results: We identified a novel lncRNA TMEM44-AS1, which was aberrantly expressed in glioma tissues, and that increased TMEM44-AS1 expression was correlated with malignant progression and poor survival for patients with glioma. Expression of TMEM44-AS1 increased the proliferation, colony formation, migration, and invasion of glioma cells. Knockdown of TMEM44-AS1 in glioma cells reduced cell proliferation, colony formation, migration and invasion, and tumor growth in a nude mouse xenograft model. Mechanistically, TMEM44-AS1 is directly bound to the SerpinB3, and sequentially activated Myc and EGR1/IL-6 signaling; Myc transcriptionally induced TMEM44-AS1 and directly bound to the promoter and super-enhancer of TMEM44-AS1, thus forming a positive feedback loop with TMEM44-AS. Further studies demonstrated that Myc interacts with MED1 regulates the super-enhancer of TMEM44-AS1. More importantly, a novel small-molecule Myc inhibitor, Myci975, alleviated TMEM44-AS1-promoted the growth of glioma cells.
Conclusions: Our study implicates a crucial role of the TMEM44-AS1-Myc axis in glioma progression and provides a possible anti-glioma therapeutic agent.
Keywords: Glioma; Myc; Super-enhancer; TMEM44-AS1; lncRNA.
© 2021. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
-
- Louis DN, Perry A, Reifenberger G, von Deimling A, Figarella-Branger D, Cavenee WK, Ohgaki H, Wiestler OD, Kleihues P, Ellison DW. The 2016 World Health Organization classification of tumors of the central nervous system: a summary. Acta Neuropathol. 2016;131(6):803–820. doi: 10.1007/s00401-016-1545-1. - DOI - PubMed
-
- F. Graus, Bruna J., Pardo J., Escudero D., Vilas D., Barcelo I., Brell M., Pascual C., Crespo J. A., Erro E., Garcia-Romero J. C., Estela J., Martino J., Garcia-Castano A., Mata E., Lema M., Gelabert M., Fuentes R., Perez P., Manzano A., Aguas J., Belenguer A., Simon A., Henriquez I., Murcia M., Vivanco R., Rojas-Marcos I., Munoz-Carmona D., Navas I., de Andres P., Mas G., Gil M., Verger E. Patterns of care and outcome for patients with glioblastoma diagnosed during 2008-2010 in Spain. Neuro Oncol.2013; 15(6):797-805. - PMC - PubMed
-
- Mikheeva SA, Mikheev AM, Petit A, Beyer R, Oxford RG, Khorasani L, Maxwell JP, Glackin CA, Wakimoto H, Gonzalez-Herrero I, Sanchez-Garcia I, Silber JR, Horner PJ, Rostomily RC. TWIST1 promotes invasion through mesenchymal change in human glioblastoma. Mol Cancer. 2010;9:194. doi: 10.1186/1476-4598-9-194. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
- 81972348/the national natural science foundation of china
- 1804h08020270/key research and development plan project of anhui province
- gxypZD2019019/college excellent youth talent support program in anhui province
- KJ2019A0267/key projects of natural science research in anhui province
- gxbjZD10/academic funding project for top talents in colleges and universities in anhui province
LinkOut - more resources
Full Text Sources
Research Materials
