

Epoxidation of juvenile hormone was a key innovation improving insect reproductive fitness
- PMID: 34697248
- PMCID: PMC8609300
- DOI: 10.1073/pnas.2109381118
Epoxidation of juvenile hormone was a key innovation improving insect reproductive fitness
Abstract
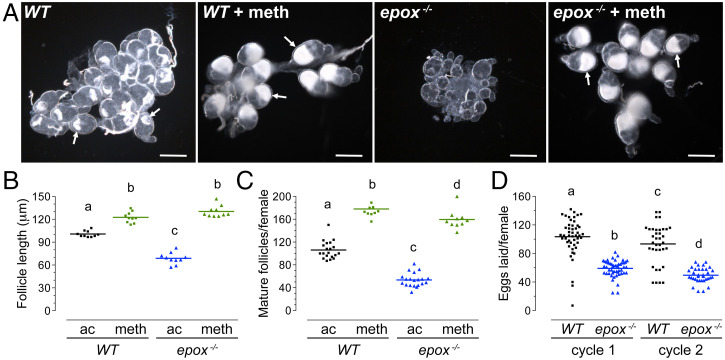
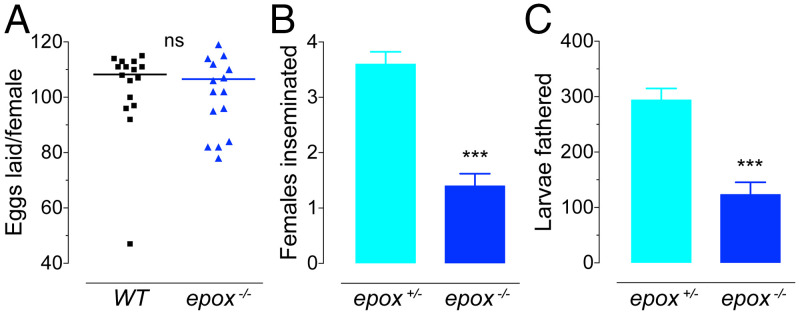
Methyl farnesoate (MF) plays hormonal regulatory roles in crustaceans. An epoxidated form of MF, known as juvenile hormone (JH), controls metamorphosis and stimulates reproduction in insects. To address the evolutionary significance of MF epoxidation, we generated mosquitoes completely lacking either of the two enzymes that catalyze the last steps of MF/JH biosynthesis and epoxidation, respectively: the JH acid methyltransferase (JHAMT) and the P450 epoxidase CYP15 (EPOX). jhamt-/- larvae lacking both MF and JH died at the onset of metamorphosis. Strikingly, epox-/- mutants, which synthesized MF but no JH, completed the entire life cycle. While epox-/- adults were fertile, the reproductive performance of both sexes was dramatically reduced. Our results suggest that although MF can substitute for the absence of JH in mosquitoes, it is with a significant fitness cost. We propose that MF can fulfill most roles of JH, but its epoxidation to JH was a key innovation providing insects with a reproductive advantage.
Keywords: Aedes aegypti; corpora allata; juvenile hormone; methyl farnesoate; reproduction.
Conflict of interest statement
The authors declare no competing interest.
Figures
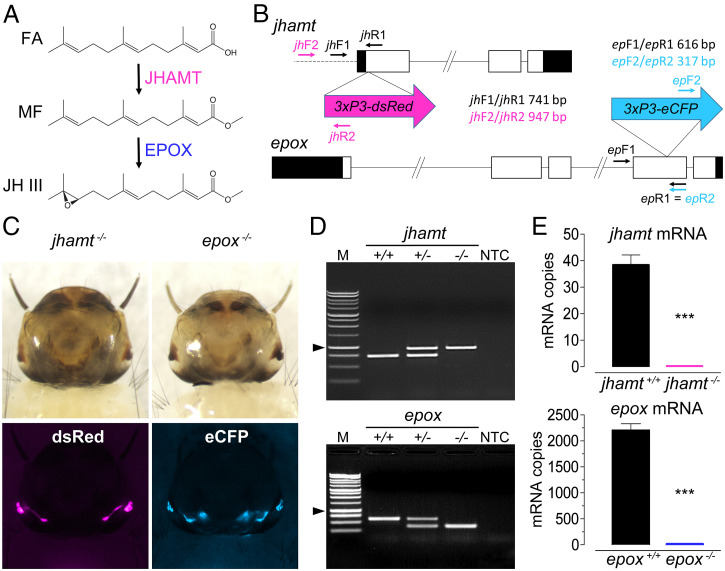
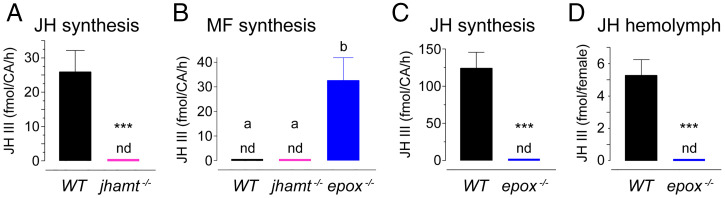
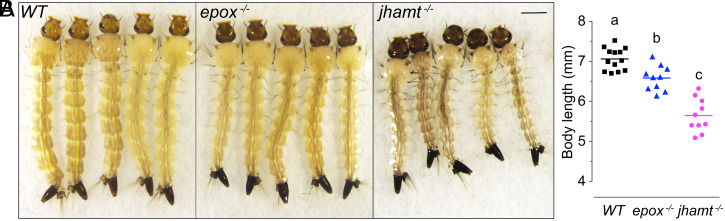
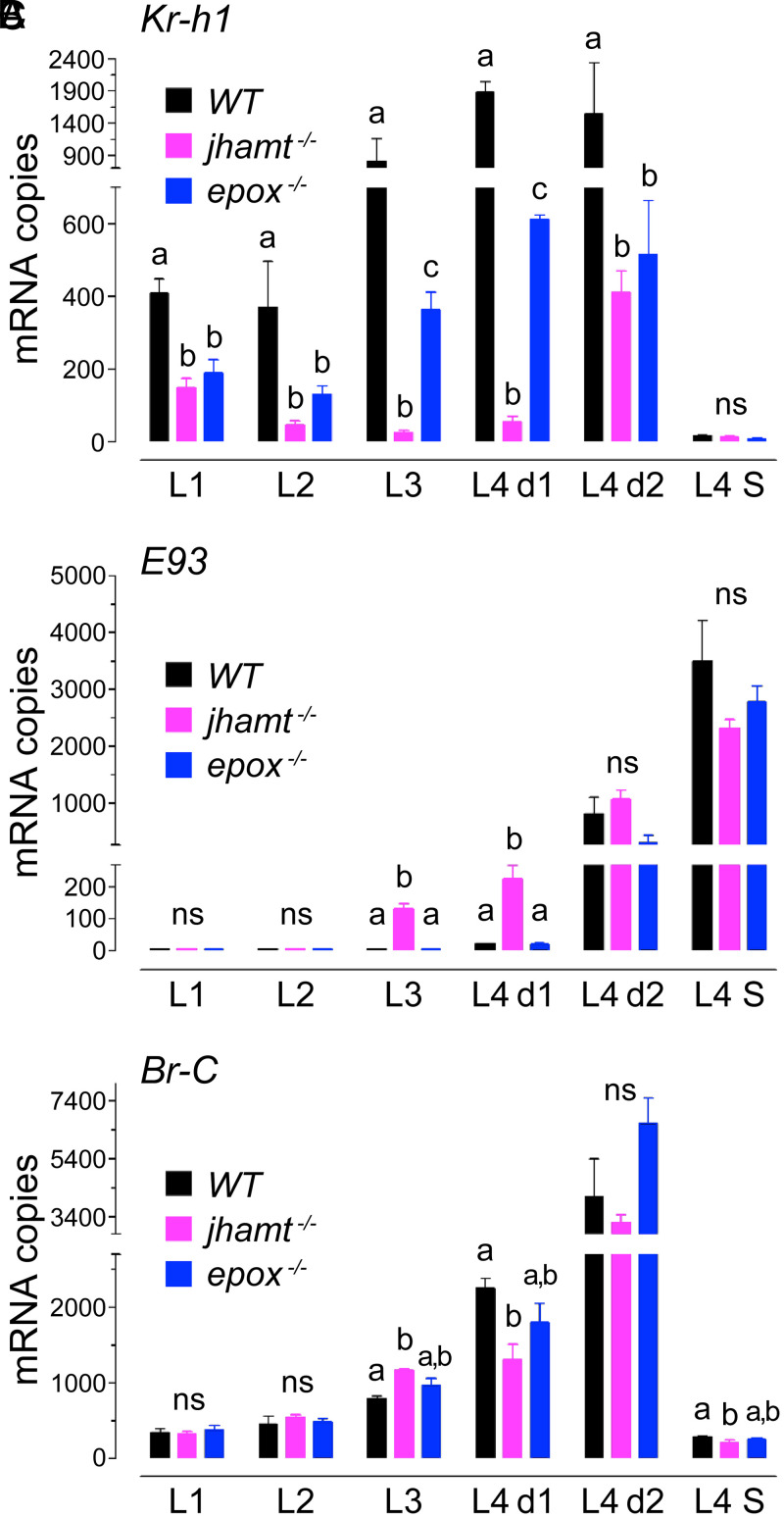
References
-
- Glenner H., Thomsen P. F., Hebsgaard M. B., Sørensen M. V., Willerslev E., Evolution. The origin of insects. Science 314, 1883–1884 (2006). - PubMed
-
- von Reumont B. M., Burmester T., “Remipedia and the evolution of hexapods” inEncyclopedia of Life Sciences (John Wiley & Sons, 2010), pp. 1–6.
-
- Giribet G., Edgecombe G. D., The phylogeny and evolutionary history of arthropods. Curr. Biol. 29, R592–R602 (2019). - PubMed
-
- Rivera-Pérez C., Clifton M. E., Noriega F. G., Jindra M., “Juvenile hormone regulation and action” in Advances in Invertebrate (Neuro)Endocrinology, Saleuddin S., Lange A. B., Orchard I., Eds. (Apple Academic Press, 2020), vol. 2, pp. 1–76.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
