

Single-stranded DNA binding proteins influence APOBEC3A substrate preference
- PMID: 34697369
- PMCID: PMC8546098
- DOI: 10.1038/s41598-021-00435-y
Single-stranded DNA binding proteins influence APOBEC3A substrate preference
Abstract
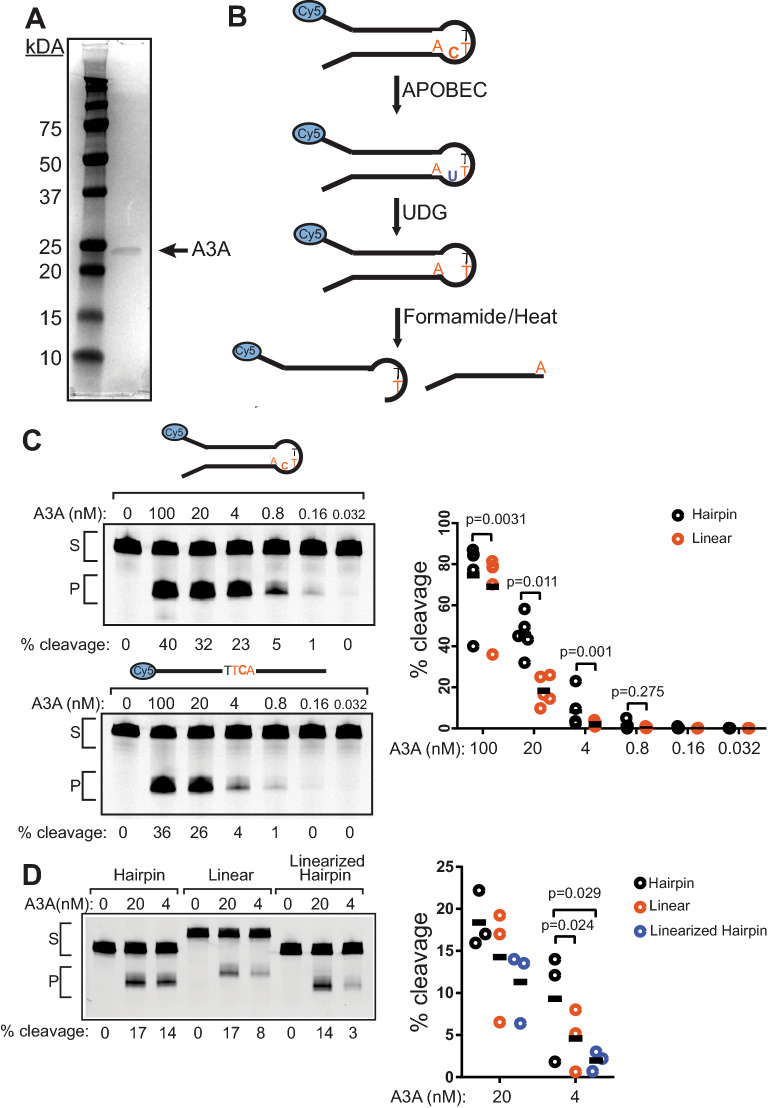
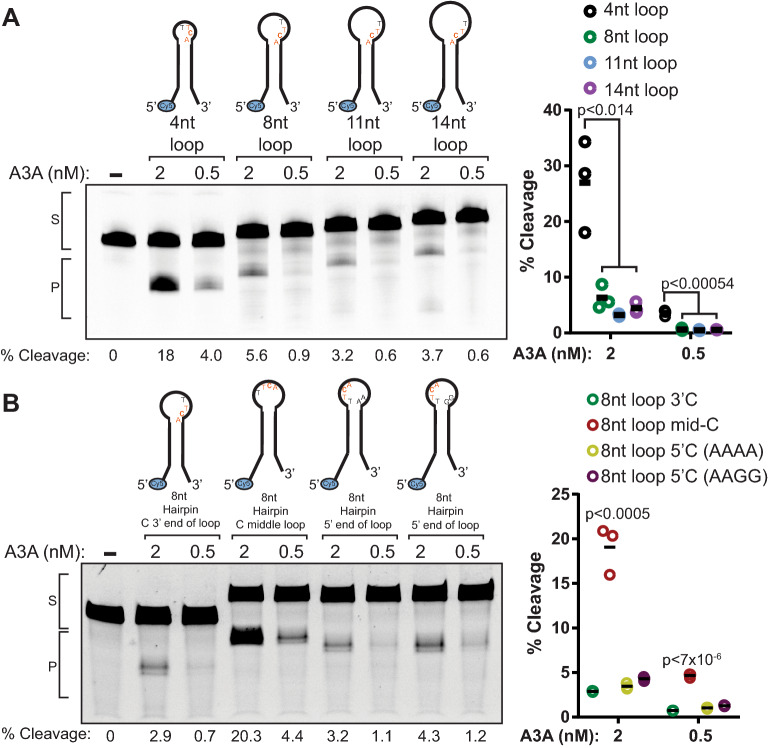
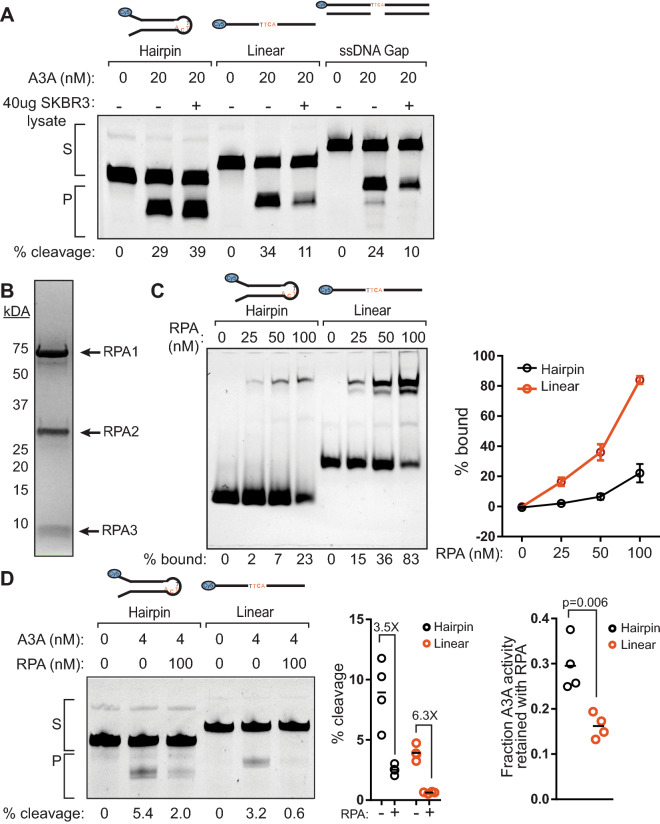
The cytidine deaminase, APOBEC3A (A3A), is a prominent source of mutations in multiple cancer types. These APOBEC-signature mutations are non-uniformly distributed across cancer genomes, associating with single-stranded (ss) DNA formed during DNA replication and hairpin-forming sequences. The biochemical and cellular factors that influence these specificities are unclear. We measured A3A's cytidine deaminase activity in vitro on substrates that model potential sources of ssDNA in the cell and found that A3A is more active on hairpins containing 4 nt ssDNA loops compared to hairpins with larger loops, bubble structures, replication fork mimics, ssDNA gaps, or linear DNA. Despite pre-bent ssDNAs being expected to fit better in the A3A active site, we determined A3A favors a 4 nt hairpin substrate only 2- to fivefold over linear ssDNA substrates. Addition of whole cell lysates or purified RPA to cytidine deaminase assays more severely reduced A3A activity on linear ssDNA (45 nt) compared to hairpin substrates. These results indicate that the large enrichment of A3A-driven mutations in hairpin-forming sequences in tumor genomes is likely driven in part by other proteins that preferentially bind longer ssDNA regions, which limit A3A's access. Furthermore, A3A activity is reduced at ssDNA associated with a stalled T7 RNA polymerase, suggesting that potential protein occlusion by RNA polymerase also limits A3A activity. These results help explain the small transcriptional strand bias for APOBEC mutation signatures in cancer genomes and the general targeting of hairpin-forming sequences in the lagging strand template during DNA replication.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


Similar articles
-
Structural determinants of human APOBEC3A enzymatic and nucleic acid binding properties.Nucleic Acids Res. 2014 Jan;42(2):1095-110. doi: 10.1093/nar/gkt945. Epub 2013 Oct 24. Nucleic Acids Res. 2014. PMID: 24163103 Free PMC article.
-
Creating RNA Specific C-to-U Editase from APOBEC3A by Separation of Its Activities on DNA and RNA Substrates.ACS Synth Biol. 2021 May 21;10(5):1106-1115. doi: 10.1021/acssynbio.0c00627. Epub 2021 May 2. ACS Synth Biol. 2021. PMID: 33938211
-
Substrate sequence selectivity of APOBEC3A implicates intra-DNA interactions.Sci Rep. 2018 May 14;8(1):7511. doi: 10.1038/s41598-018-25881-z. Sci Rep. 2018. PMID: 29760455 Free PMC article.
-
Functions and regulation of the APOBEC family of proteins.Semin Cell Dev Biol. 2012 May;23(3):258-68. doi: 10.1016/j.semcdb.2011.10.004. Epub 2011 Oct 6. Semin Cell Dev Biol. 2012. PMID: 22001110 Free PMC article. Review.
-
Breaking bad: The mutagenic effect of DNA repair.DNA Repair (Amst). 2015 Aug;32:43-51. doi: 10.1016/j.dnarep.2015.04.012. Epub 2015 May 1. DNA Repair (Amst). 2015. PMID: 26073774 Free PMC article. Review.
Cited by
-
Addressing the benefits of inhibiting APOBEC3-dependent mutagenesis in cancer.Nat Genet. 2022 Nov;54(11):1599-1608. doi: 10.1038/s41588-022-01196-8. Epub 2022 Oct 24. Nat Genet. 2022. PMID: 36280735 Free PMC article. Review.
-
Competition for DNA binding between the genome protector replication protein A and the genome modifying APOBEC3 single-stranded DNA deaminases.Nucleic Acids Res. 2022 Nov 28;50(21):12039-12057. doi: 10.1093/nar/gkac1121. Nucleic Acids Res. 2022. PMID: 36444883 Free PMC article.
-
APOBEC3: Friend or Foe in Human Papillomavirus Infection and Oncogenesis?Annu Rev Virol. 2022 Sep 29;9(1):375-395. doi: 10.1146/annurev-virology-092920-030354. Epub 2022 Jun 7. Annu Rev Virol. 2022. PMID: 35671565 Free PMC article. Review.
-
Hypomorphic mutation in the large subunit of replication protein A affects mutagenesis by human APOBEC cytidine deaminases in yeast.G3 (Bethesda). 2024 Oct 7;14(10):jkae196. doi: 10.1093/g3journal/jkae196. G3 (Bethesda). 2024. PMID: 39150943 Free PMC article.
-
Mesoscale DNA features impact APOBEC3A and APOBEC3B deaminase activity and shape tumor mutational landscapes.Nat Commun. 2024 Mar 18;15(1):2370. doi: 10.1038/s41467-024-45909-5. Nat Commun. 2024. PMID: 38499542 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
