

Context-dependent representations of movement in Drosophila dopaminergic reinforcement pathways
- PMID: 34697455
- PMCID: PMC8556349
- DOI: 10.1038/s41593-021-00929-y
Context-dependent representations of movement in Drosophila dopaminergic reinforcement pathways
Abstract
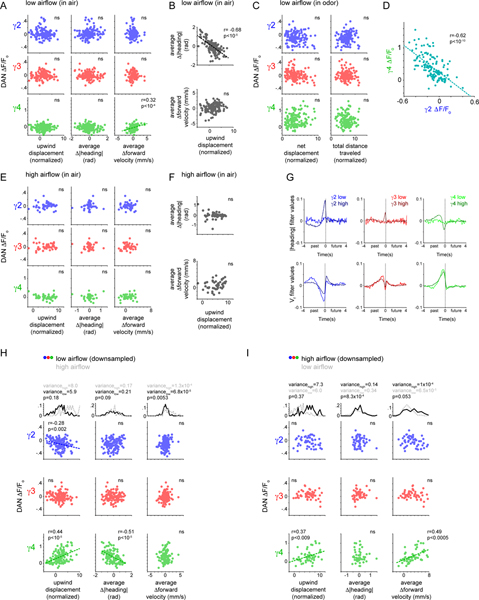
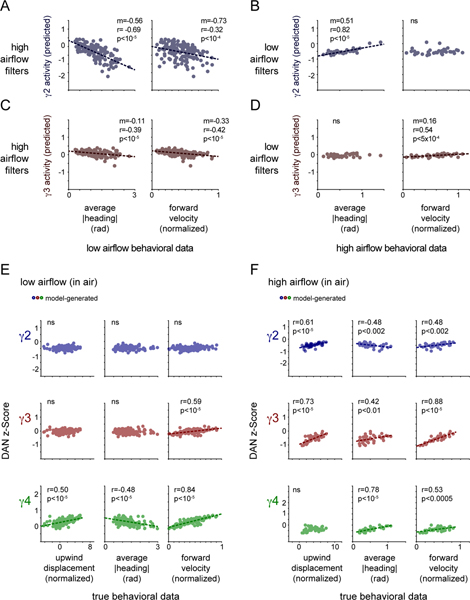
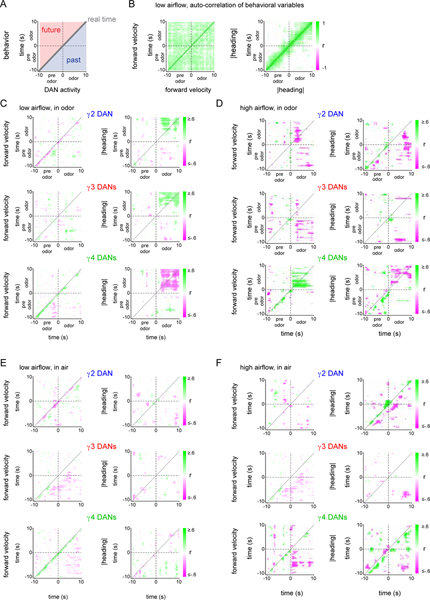
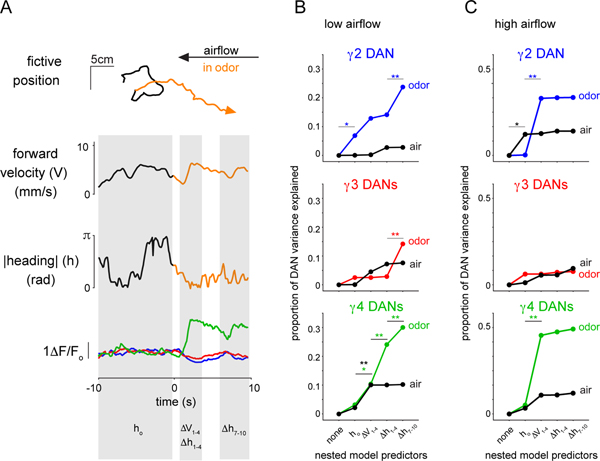
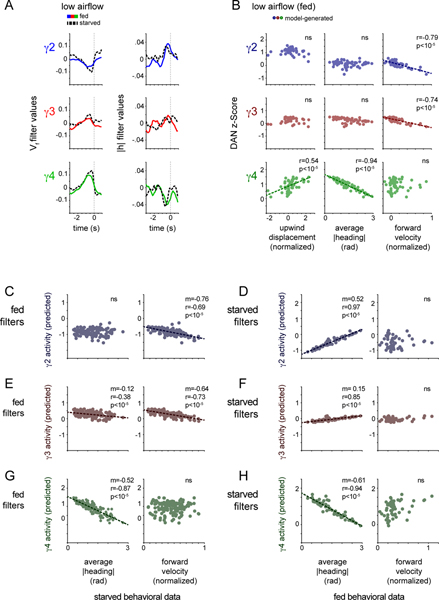
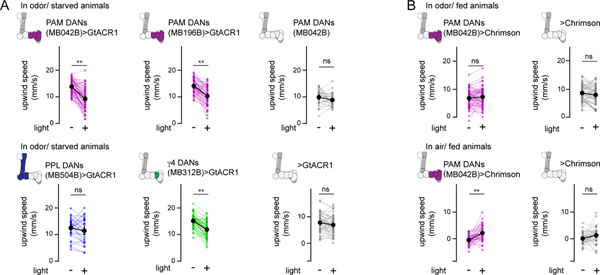
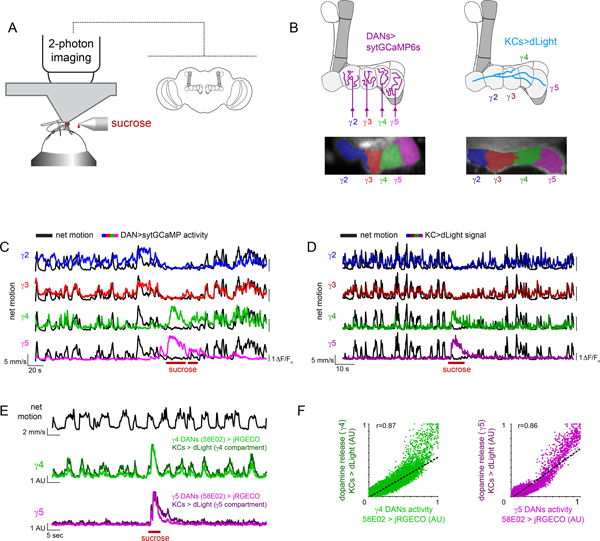
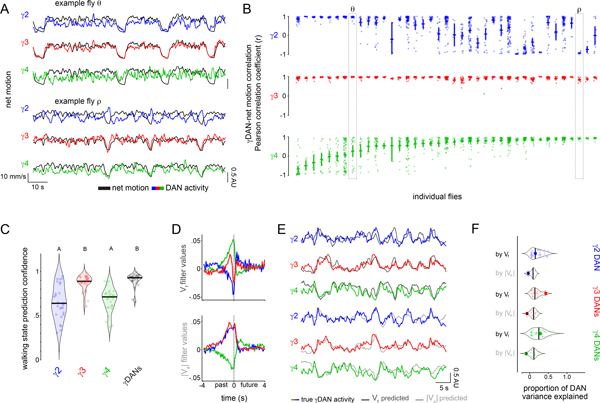
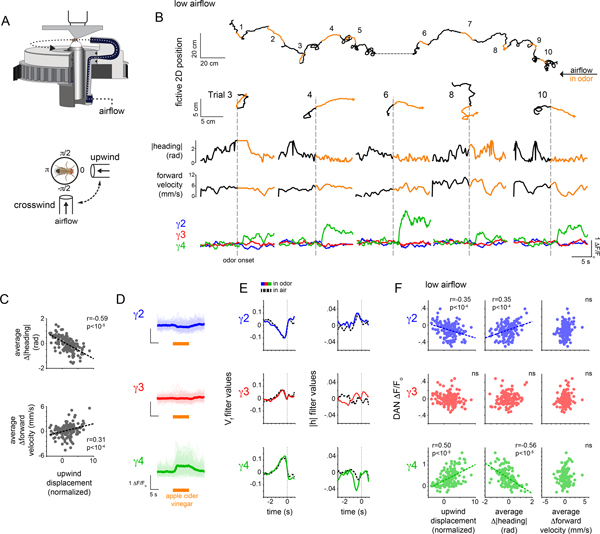
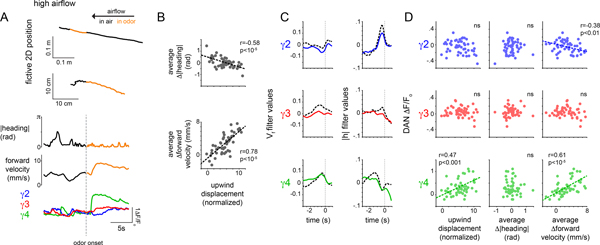
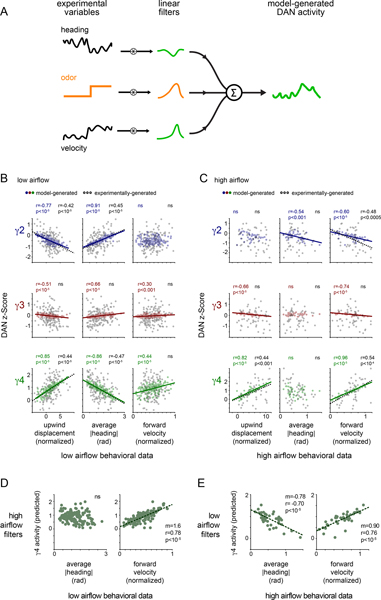
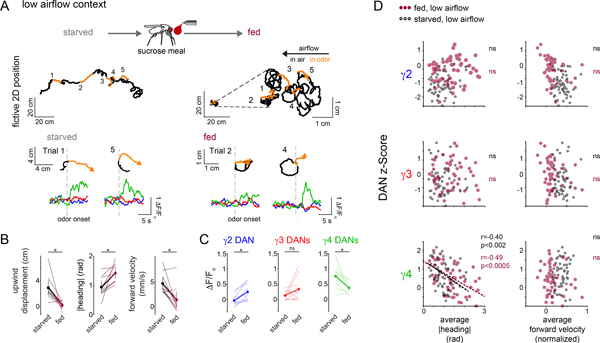
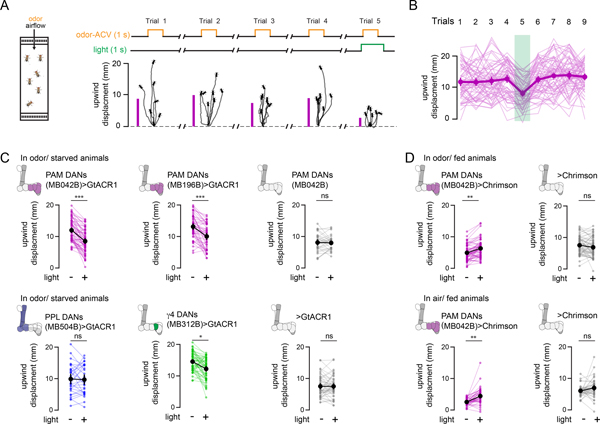
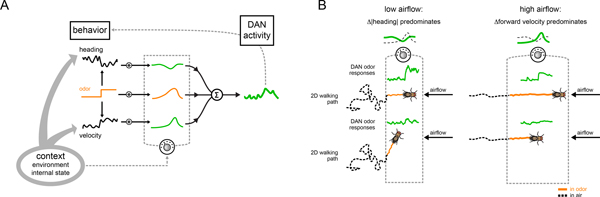
Dopamine plays a central role in motivating and modifying behavior, serving to invigorate current behavioral performance and guide future actions through learning. Here we examine how this single neuromodulator can contribute to such diverse forms of behavioral modulation. By recording from the dopaminergic reinforcement pathways of the Drosophila mushroom body during active odor navigation, we reveal how their ongoing motor-associated activity relates to goal-directed behavior. We found that dopaminergic neurons correlate with different behavioral variables depending on the specific navigational strategy of an animal, such that the activity of these neurons preferentially reflects the actions most relevant to odor pursuit. Furthermore, we show that these motor correlates are translated to ongoing dopamine release, and acutely perturbing dopaminergic signaling alters the strength of odor tracking. Context-dependent representations of movement and reinforcement cues are thus multiplexed within the mushroom body dopaminergic pathways, enabling them to coordinately influence both ongoing and future behavior.
© 2021. The Author(s), under exclusive licence to Springer Nature America, Inc.
Conflict of interest statement
Competing Interests Statement
The authors declare no competing interests.
Figures
References
-
- Bargmann CI & Marder E From the connectome to brain function. Nat. Methods 10, 483–490 (2013). - PubMed
-
- Schultz W, Dayan P, & Montague PR A neural substrate of prediction and reward. Science 275, 1593–1599 (1997). - PubMed
-
- Da Silva JA, Tecuapetla F, Paixão V & Costa RM Dopamine neuron activity before action initiation gates and invigorates future movements. Nature 554, 244–248 (2018). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
