

Dnmt3a-mutated clonal hematopoiesis promotes osteoporosis
- PMID: 34698806
- PMCID: PMC8552148
- DOI: 10.1084/jem.20211872
Dnmt3a-mutated clonal hematopoiesis promotes osteoporosis
Abstract
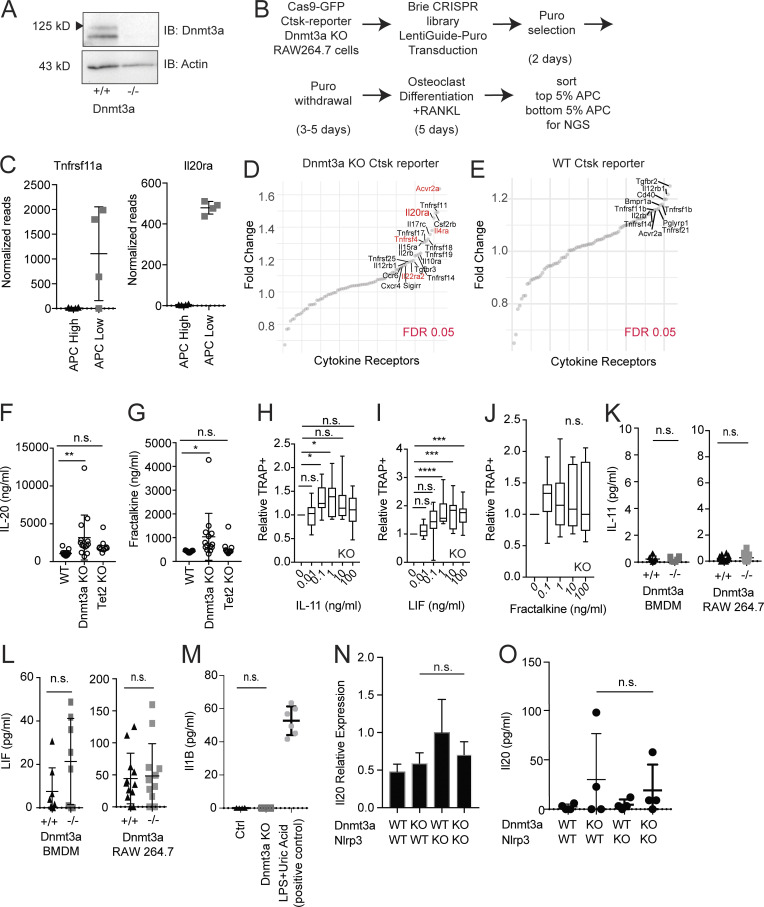
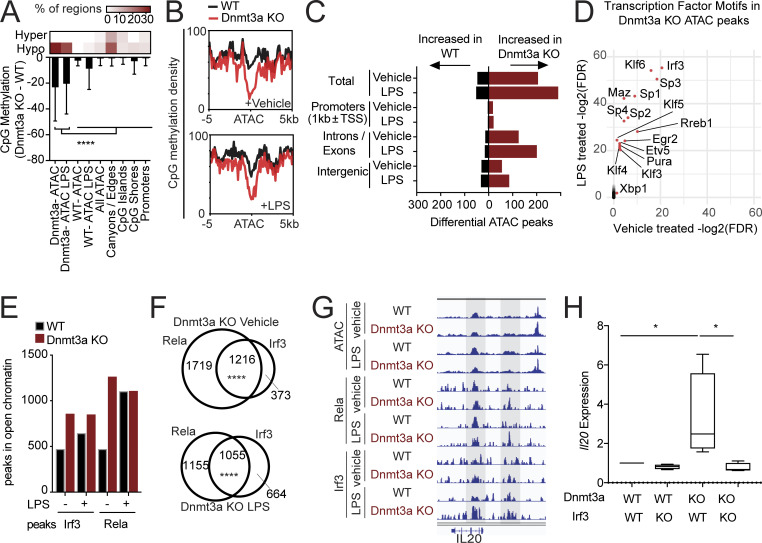
Osteoporosis is caused by an imbalance of osteoclasts and osteoblasts, occurring in close proximity to hematopoietic cells in the bone marrow. Recurrent somatic mutations that lead to an expanded population of mutant blood cells is termed clonal hematopoiesis of indeterminate potential (CHIP). Analyzing exome sequencing data from the UK Biobank, we found CHIP to be associated with increased incident osteoporosis diagnoses and decreased bone mineral density. In murine models, hematopoietic-specific mutations in Dnmt3a, the most commonly mutated gene in CHIP, decreased bone mass via increased osteoclastogenesis. Dnmt3a-/- demethylation opened chromatin and altered activity of inflammatory transcription factors. Bone loss was driven by proinflammatory cytokines, including Irf3-NF-κB-mediated IL-20 expression from Dnmt3a mutant macrophages. Increased osteoclastogenesis due to the Dnmt3a mutations was ameliorated by alendronate or IL-20 neutralization. These results demonstrate a novel source of osteoporosis-inducing inflammation.
© 2021 Kim et al.
Conflict of interest statement
Disclosures: A. Niroula reported grants from the Knut and Alice Wallenberg Foundation outside the submitted work. A. Bick reported personal fees from Foresite Labs outside the submitted work. J. Pirruccello reported personal fees from Maze Therapeutics outside the submitted work. M. Agrawal reported personal fees from German Accelerator Life Sciences outside the submitted work, and is the co-founder of and holds equity in iuvando Health. None of these are related to the submitted work. D.P. Kiel reported grants from Amgen, Radius Health, and Solarea Bio; and "other" from Solarea Bio, Pfizer, and Wolters Kluwer outside the submitted work. J.B. Richards reported personal fees from GlaxoSmithKline and Deerfield Capital; grants from Biogen and Eli Lilly; and non-financial support from 5 Prime Sciences outside the submitted work. J.B. Richards has served as an advisor to GlaxoSmithKline and Deerfield Capital. His institution has received investigator-initiated grant funding from Eli Lilly, GlaxoSmithKline and Biogen for projects unrelated to this research. He is the founder of 5 Prime Sciences. M.N. Wein reported grants from Radius Health and Galapagos NV, and "other" from Relation Therapeutics outside the submitted work. S. Jaiswal reported personal fees from AVRO Bio, Novartis, Genentech, and Foresite Labs outside the submitted work. P. Natarajan reported grants from Apple, AstraZeneca, Boston Scientific, and Novartis; personal fees from Apple, AstraZeneca, Genentech, Novartis, Blackstone Life Sciences, and Foresite Labs; and "other" from Vertex outside the submitted work. B.L. Ebert reported grants from Celgene, Novartis, Deerfield, and Calico; and personal fees from Exo Therapeutics, Skyhawk Therapeutics, Neomorph Therapeutics, and TenSixteen Bio outside the submitted work. No other disclosures were reported.
Figures






Comment in
-
Bone marrow runs the (bone) show.J Exp Med. 2021 Dec 6;218(12):e20211996. doi: 10.1084/jem.20211996. Epub 2021 Oct 27. J Exp Med. 2021. PMID: 34705037 Free PMC article.
-
Somatic mutations linked to osteoporosis.Nat Rev Rheumatol. 2022 Jan;18(1):4. doi: 10.1038/s41584-021-00727-7. Nat Rev Rheumatol. 2022. PMID: 34837067 No abstract available.
References
-
- Ambrosi, T.H., Scialdone A., Graja A., Gohlke S., Jank A.M., Bocian C., Woelk L., Fan H., Logan D.W., Schürmann A., et al. 2017. Adipocyte Accumulation in the Bone Marrow during Obesity and Aging Impairs Stem Cell-Based Hematopoietic and Bone Regeneration. Cell Stem Cell. 20:771–784.e6. 10.1016/j.stem.2017.02.009 - DOI - PMC - PubMed
-
- Benjamin, D., Sato T., Cibulskis K., Getz G., Stewart C., and Lichtenstein L.. 2019. Calling Somatic SNVs and Indels with Mutect2. bioRxiv. 10.1101/861054 (Preprint posted December 2, 2019) - DOI
-
- Bieghs, V., Van Gorp P.J., Wouters K., Hendrikx T., Gijbels M.J., van Bilsen M., Bakker J., Binder C.J., Lütjohann D., Staels B., et al. 2012. LDL receptor knock-out mice are a physiological model particularly vulnerable to study the onset of inflammation in non-alcoholic fatty liver disease. PLoS One. 7:e30668. 10.1371/journal.pone.0030668 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R35 CA253125/CA/NCI NIH HHS/United States
- P50 CA206963/CA/NCI NIH HHS/United States
- MC_QA137853/MRC_/Medical Research Council/United Kingdom
- R01 AR041398/AR/NIAMS NIH HHS/United States
- R21AR077768/AR/NIAMS NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- P01 CA108631/CA/NCI NIH HHS/United States
- MC_UU_12013/4/MRC_/Medical Research Council/United Kingdom
- DP2 HL157540/HL/NHLBI NIH HHS/United States
- R01HL082945/NH/NIH HHS/United States
- R01DK116716/NH/NIH HHS/United States
- R01 DK116716/DK/NIDDK NIH HHS/United States
- R01 HL082945/HL/NHLBI NIH HHS/United States
- DP2-HL157540/NH/NIH HHS/United States
- T32 HL116324/HL/NHLBI NIH HHS/United States
- P01 CA066996/CA/NCI NIH HHS/United States
- PST-35-21/DRCRF/Damon Runyon Cancer Research Foundation/United States
- R01AR041398/AR/NIAMS NIH HHS/United States
- R21 AR077768/AR/NIAMS NIH HHS/United States
- MC_PC_17228/MRC_/Medical Research Council/United Kingdom
- 5T32HL116324/HL/NHLBI NIH HHS/United States
- C18281/A29019/CRUK_/Cancer Research UK/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
