

Murine SEC24D can substitute functionally for SEC24C during embryonic development
- PMID: 34702932
- PMCID: PMC8548507
- DOI: 10.1038/s41598-021-00579-x
Murine SEC24D can substitute functionally for SEC24C during embryonic development
Abstract
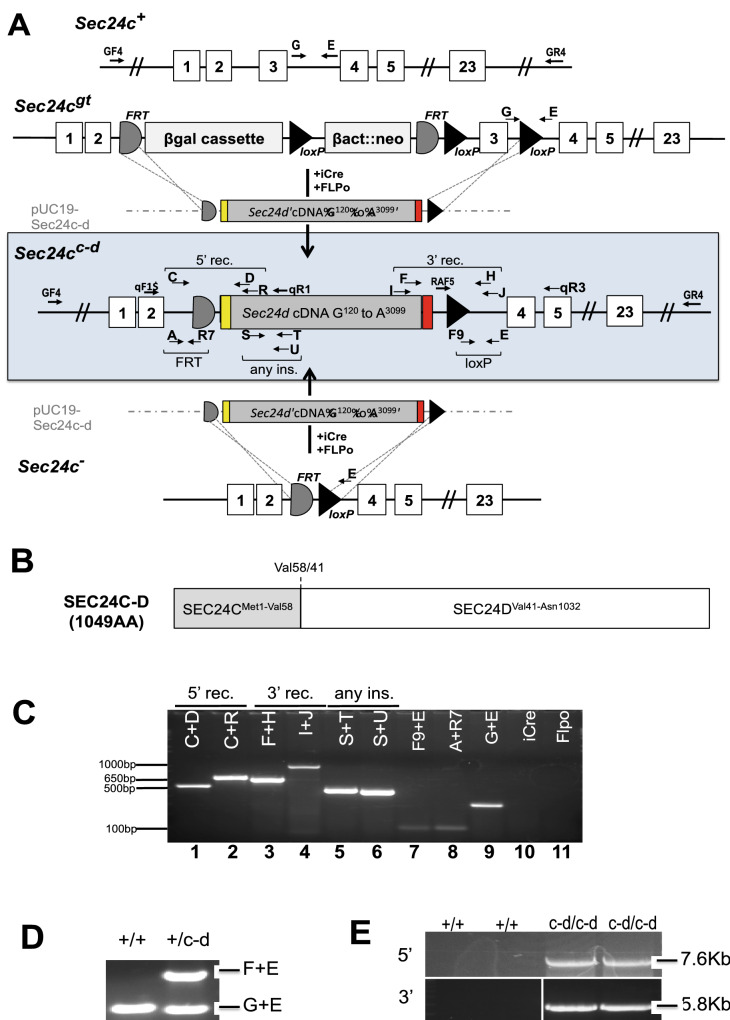
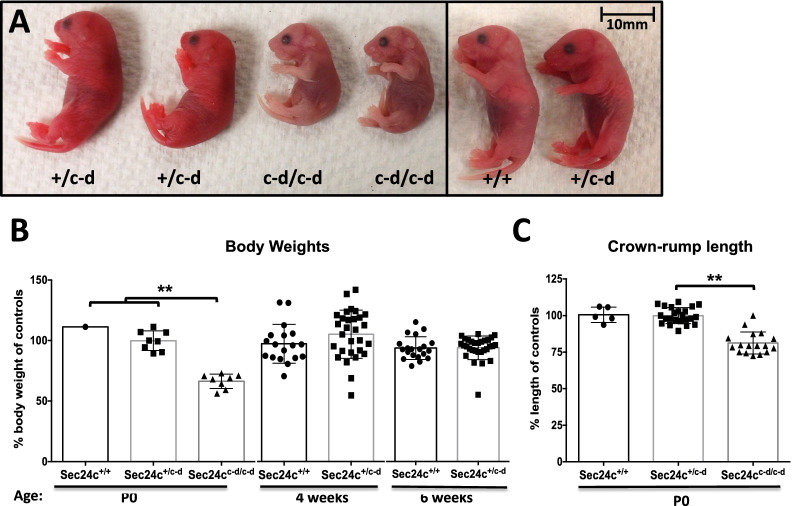
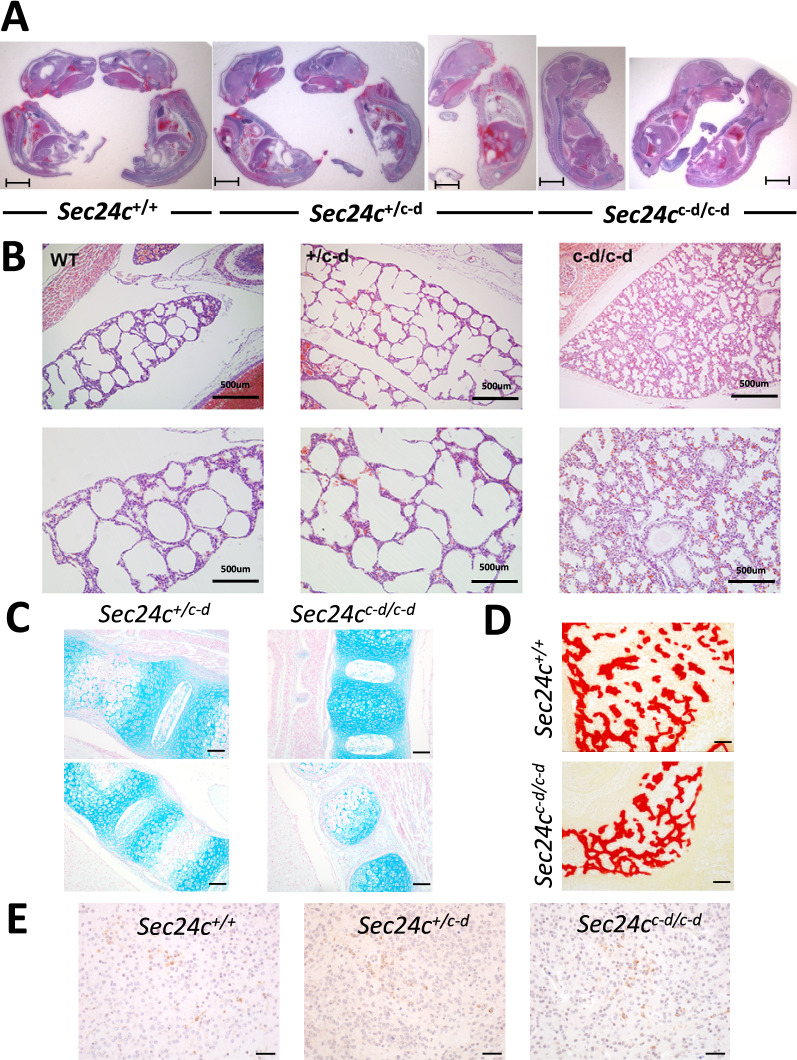
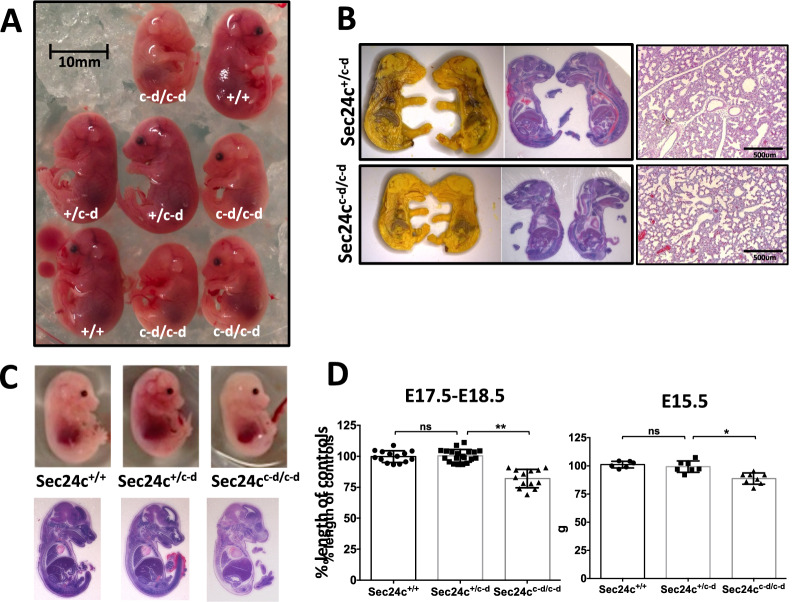
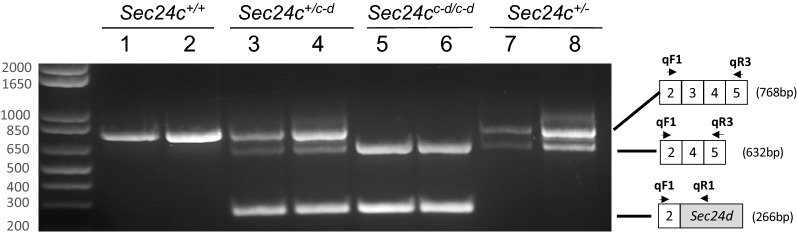
The COPII component SEC24 mediates the recruitment of transmembrane cargos or cargo adaptors into newly forming COPII vesicles on the ER membrane. Mammalian genomes encode four Sec24 paralogs (Sec24a-d), with two subfamilies based on sequence homology (SEC24A/B and C/D), though little is known about their comparative functions and cargo-specificities. Complete deficiency for Sec24d results in very early embryonic lethality in mice (before the 8 cell stage), with later embryonic lethality (E7.5) observed in Sec24c null mice. To test the potential overlap in function between SEC24C/D, we employed dual recombinase mediated cassette exchange to generate a Sec24cc-d allele, in which the C-terminal 90% of SEC24C has been replaced by SEC24D coding sequence. In contrast to the embryonic lethality at E7.5 of SEC24C-deficiency, Sec24cc-d/c-d pups survive to term, though dying shortly after birth. Sec24cc-d/c-d pups are smaller in size, but exhibit no other obvious developmental abnormality by pathologic evaluation. These results suggest that tissue-specific and/or stage-specific expression of the Sec24c/d genes rather than differences in cargo export function explain the early embryonic requirements for SEC24C and SEC24D.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Mammalian COPII coat component SEC24C is required for embryonic development in mice.J Biol Chem. 2014 Jul 25;289(30):20858-70. doi: 10.1074/jbc.M114.566687. J Biol Chem. 2014. PMID: 24876386 Free PMC article.
-
The COPII cargo adapter SEC24C is essential for neuronal homeostasis.J Clin Invest. 2018 Aug 1;128(8):3319-3332. doi: 10.1172/JCI98194. Epub 2018 Jun 25. J Clin Invest. 2018. PMID: 29939162 Free PMC article.
-
Collagen has a unique SEC24 preference for efficient export from the endoplasmic reticulum.Traffic. 2022 Jan;23(1):81-93. doi: 10.1111/tra.12826. Epub 2021 Nov 22. Traffic. 2022. PMID: 34761479 Free PMC article.
-
Consequences of mutations in the genes of the ER export machinery COPII in vertebrates.Cell Stress Chaperones. 2020 Mar;25(2):199-209. doi: 10.1007/s12192-019-01062-3. Epub 2020 Jan 22. Cell Stress Chaperones. 2020. PMID: 31970693 Free PMC article. Review.
-
The pathway of collagen secretion.Annu Rev Cell Dev Biol. 2015;31:109-24. doi: 10.1146/annurev-cellbio-100913-013002. Epub 2015 Sep 29. Annu Rev Cell Dev Biol. 2015. PMID: 26422332 Review.
Cited by
-
SEC24D depletion induces osteogenic differentiation deficiency by inactivating the ATF6/TGF-β/Runx2 regulatory loop.Commun Biol. 2025 May 15;8(1):758. doi: 10.1038/s42003-025-08175-9. Commun Biol. 2025. PMID: 40374976 Free PMC article.
-
Cargo selection in endoplasmic reticulum-to-Golgi transport and relevant diseases.J Clin Invest. 2023 Jan 3;133(1):e163838. doi: 10.1172/JCI163838. J Clin Invest. 2023. PMID: 36594468 Free PMC article. Review.
-
ER-to-Golgi Trafficking and Its Implication in Neurological Diseases.Cells. 2020 Feb 11;9(2):408. doi: 10.3390/cells9020408. Cells. 2020. PMID: 32053905 Free PMC article. Review.
-
Functional overlap between the mammalian Sar1a and Sar1b paralogs in vivo.Proc Natl Acad Sci U S A. 2024 May 7;121(19):e2322164121. doi: 10.1073/pnas.2322164121. Epub 2024 Apr 30. Proc Natl Acad Sci U S A. 2024. PMID: 38687799 Free PMC article.
-
Functional overlap between the mammalian Sar1a and Sar1b paralogs in vivo.bioRxiv [Preprint]. 2024 Feb 29:2024.02.27.582310. doi: 10.1101/2024.02.27.582310. bioRxiv. 2024. Update in: Proc Natl Acad Sci U S A. 2024 May 7;121(19):e2322164121. doi: 10.1073/pnas.2322164121. PMID: 38463989 Free PMC article. Updated. Preprint.
References
Publication types
MeSH terms
Substances
Grants and funding
- P01 HL057346/HL/NHLBI NIH HHS/United States
- T32-GM007315/GM/NIGMS NIH HHS/United States
- R35 HL135793/HL/NHLBI NIH HHS/United States
- P30 CA046592/CA/NCI NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- R01 HL039693/HL/NHLBI NIH HHS/United States
- CA46592/CA/NCI NIH HHS/United States
- P30CA046592/CA/NCI NIH HHS/United States
- RO1HL148333/HL/NHLBI NIH HHS/United States
- R01 HL157062/HL/NHLBI NIH HHS/United States
- R01 HL148333/HL/NHLBI NIH HHS/United States
- T32 GM007315/GM/NIGMS NIH HHS/United States
- AR20557/AR/NIAMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
