

β2-adrenergic receptor signaling regulates metabolic pathways critical to myeloid-derived suppressor cell function within the TME
- PMID: 34706232
- PMCID: PMC8601406
- DOI: 10.1016/j.celrep.2021.109883
β2-adrenergic receptor signaling regulates metabolic pathways critical to myeloid-derived suppressor cell function within the TME
Abstract
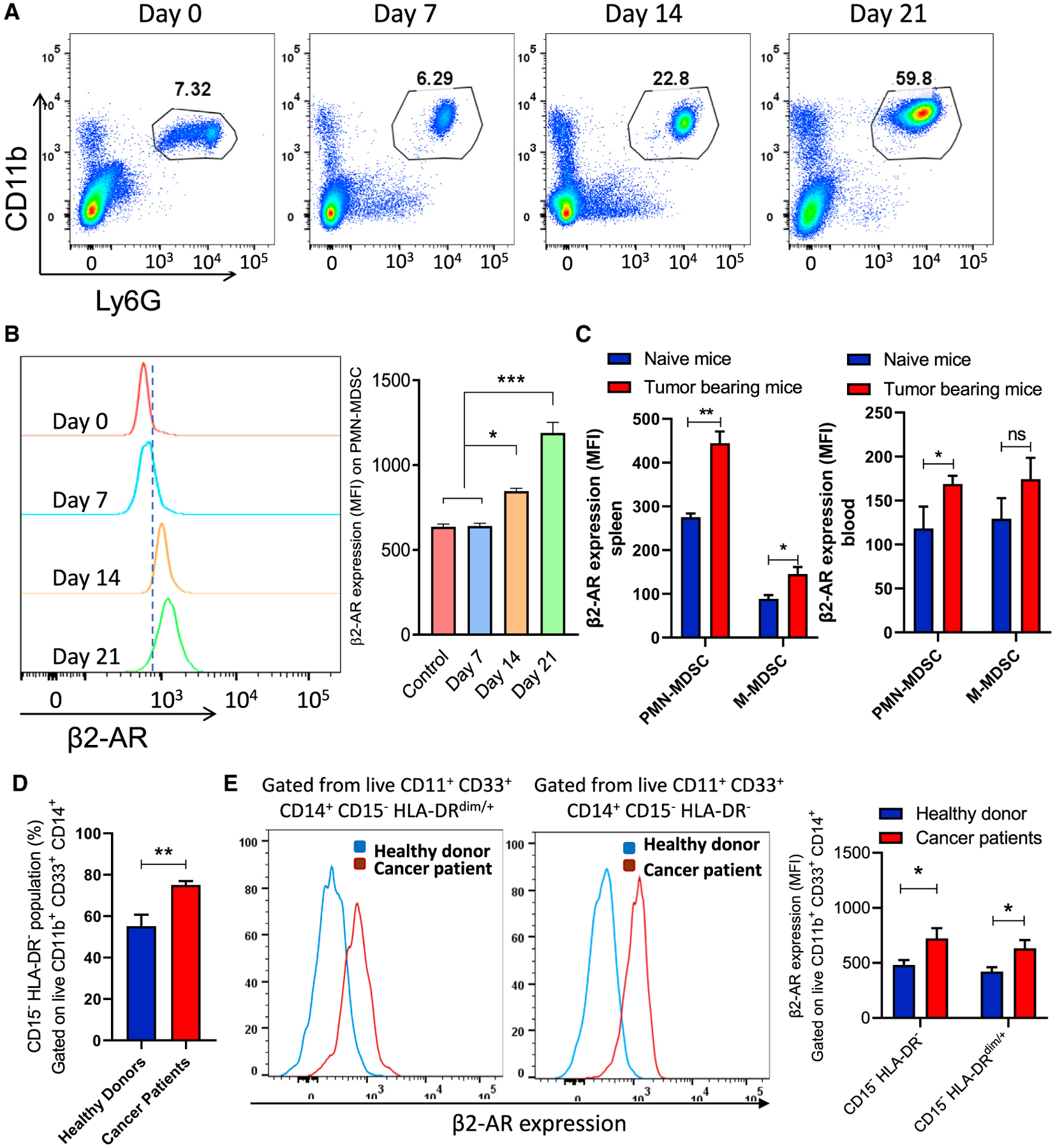
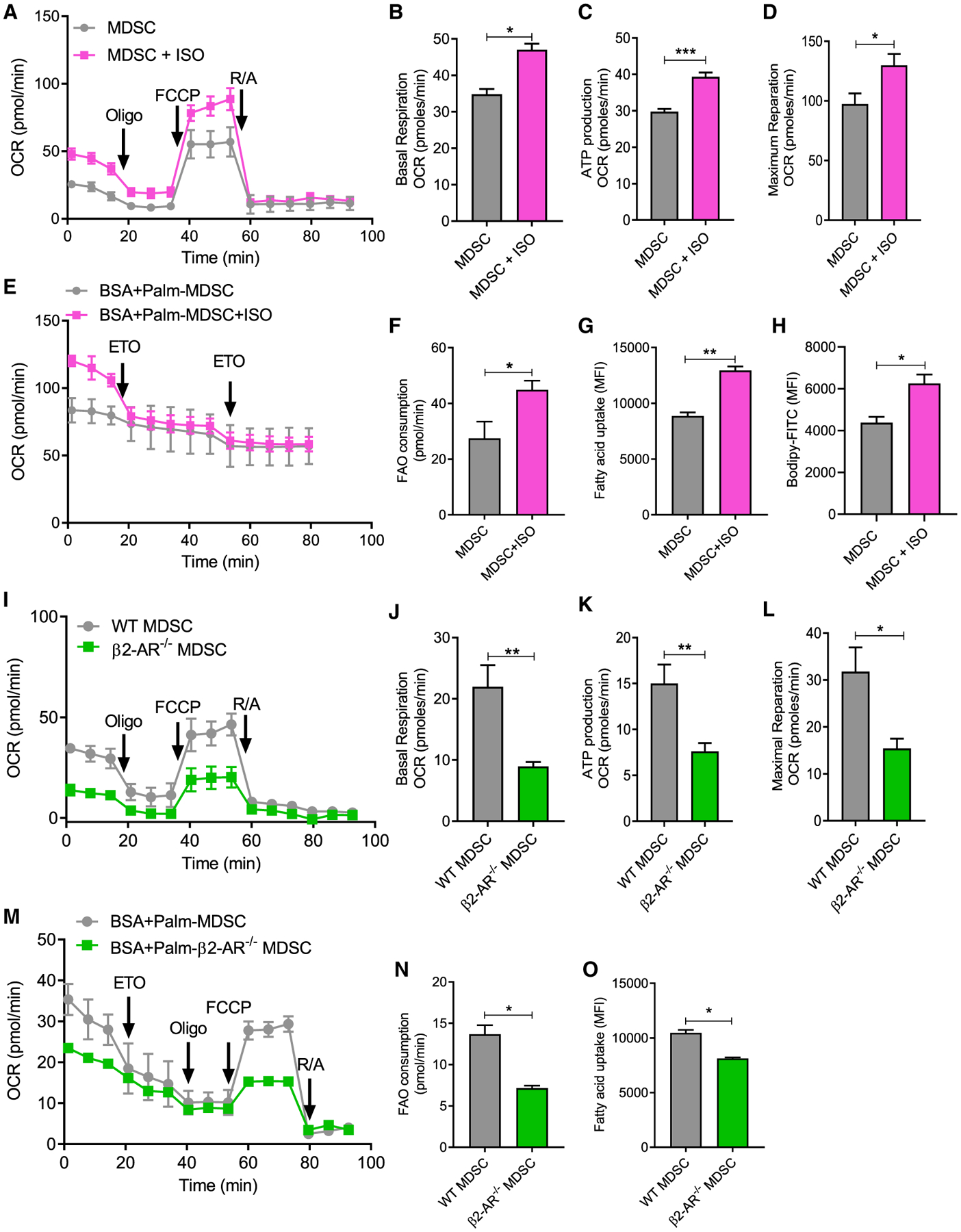
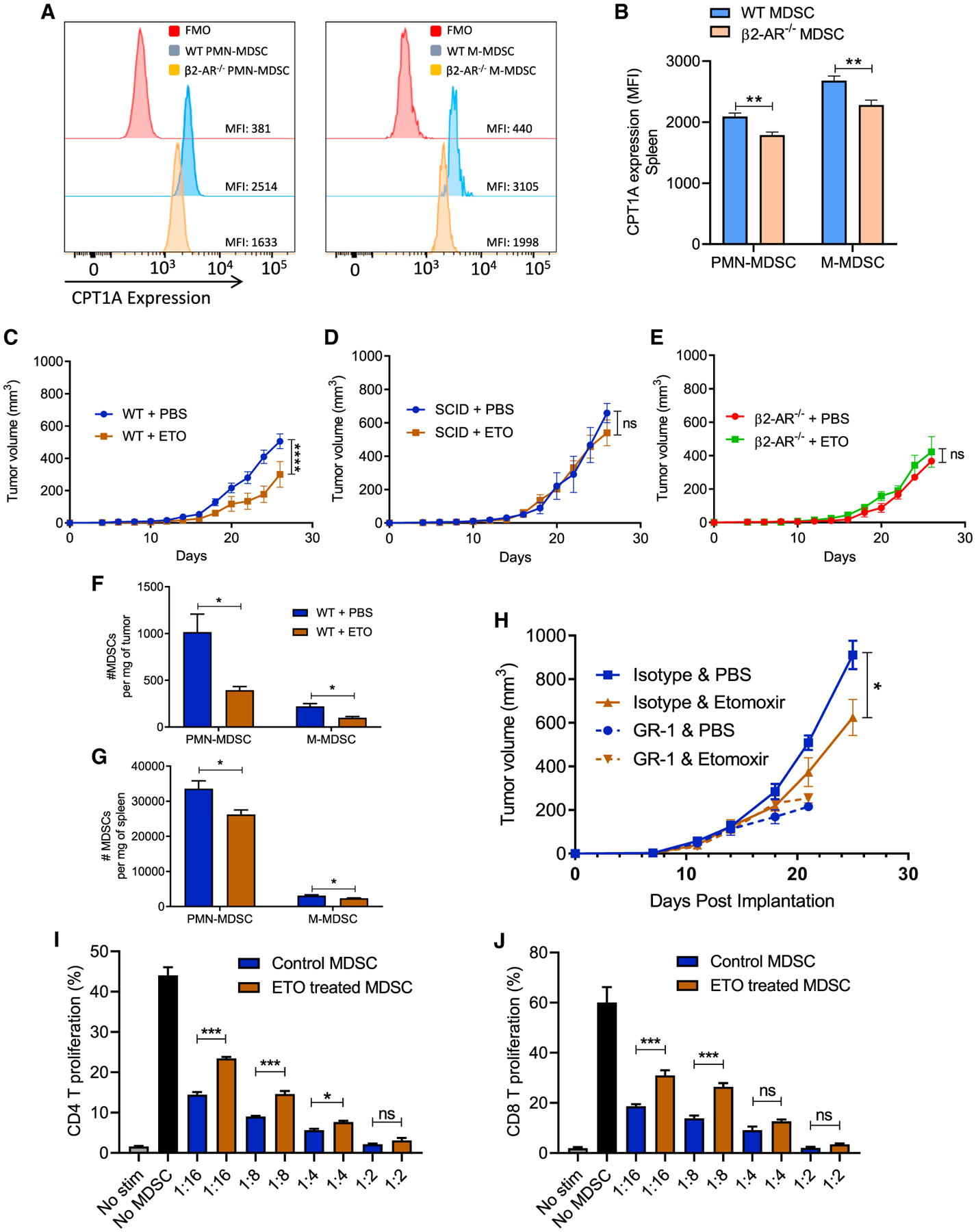
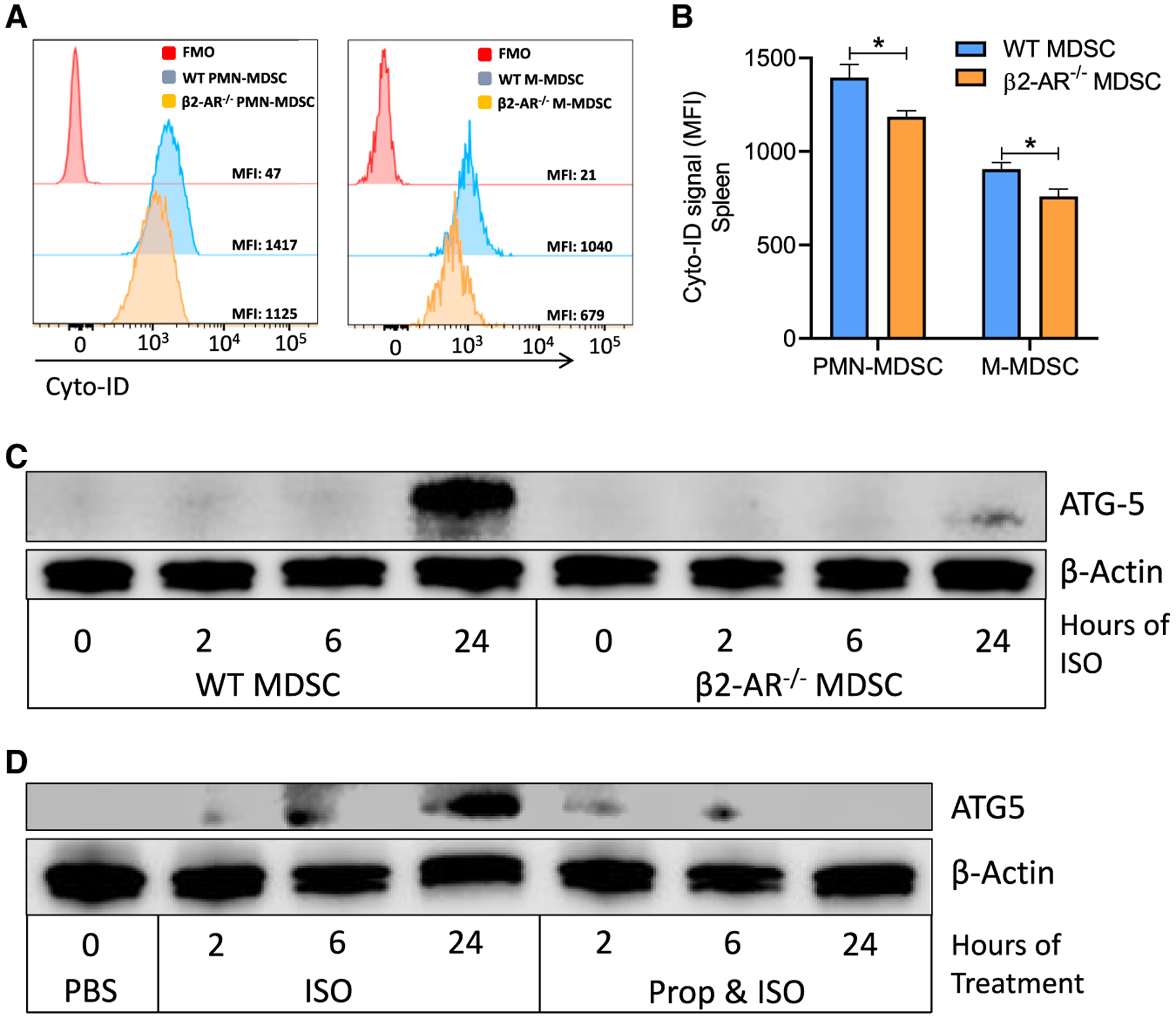
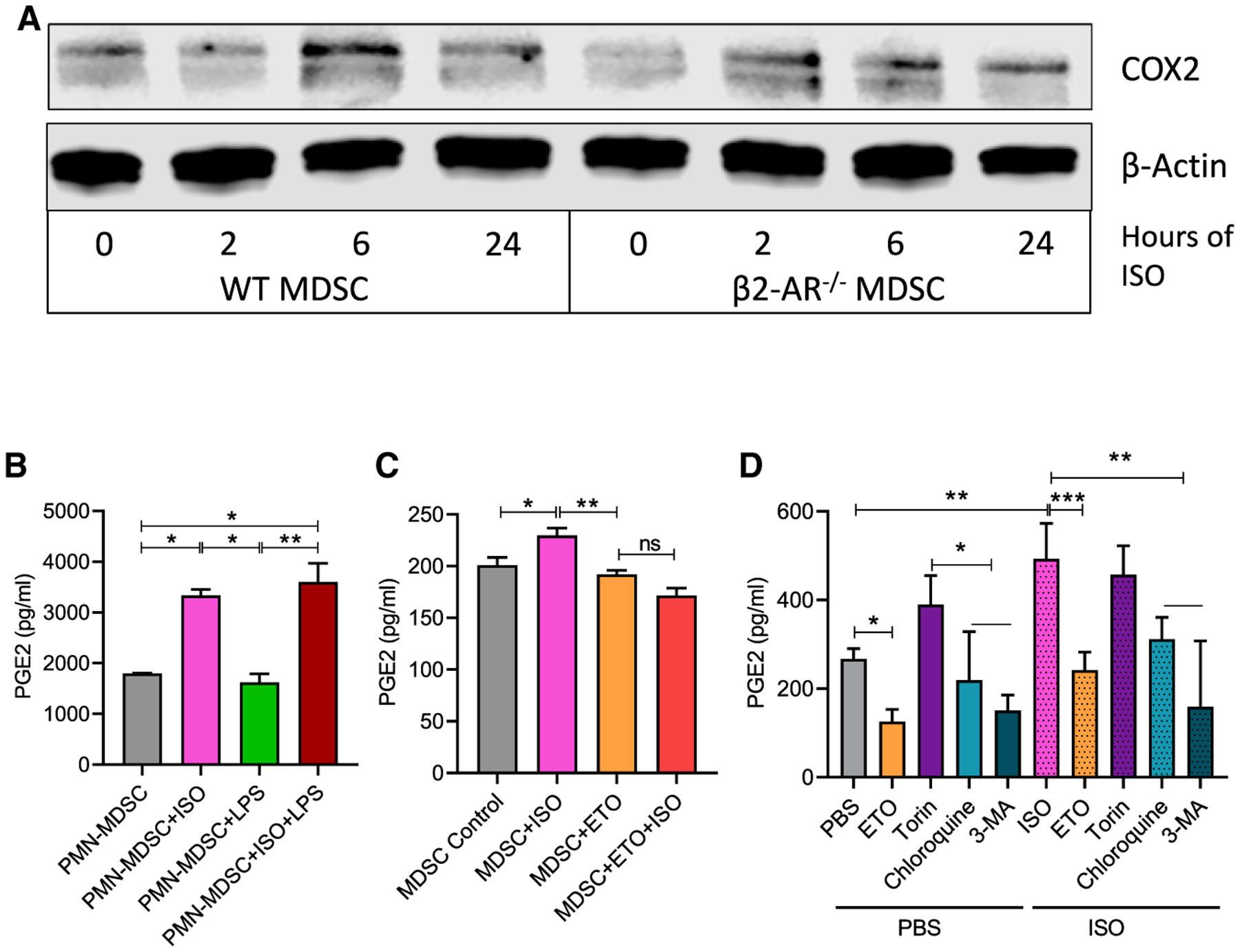
Myeloid-derived suppressor cells (MDSCs) impede antitumor immunity; however, the precise mechanisms that regulate their suppressive function remain unresolved. Identifying these mechanisms could lead to therapeutic interventions to boost cancer immunotherapy efficacy. Here, we reveal that β2 adrenergic receptor (β2-AR) expression on MDSCs increases with tumor growth and that the β2-AR stress pathway drives the immune suppressive activity of MDSCs by altering their metabolism. We show that β2-AR signaling decreases glycolysis and increases oxidative phosphorylation and fatty acid oxidation (FAO). It also increases expression of the fatty acid transporter CPT1A, which is necessary for the FAO-mediated immunosuppressive function of MDSCs. Moreover, we show that β2-AR signaling increases autophagy and activates the arachidonic acid cycle, both required for increasing the release of the immunosuppressive mediator, PGE2. Our data reveal that β2-AR signaling triggered by stress is an important physiological regulator of key metabolic pathways in MDSCs, driving their immunosuppressive function.
Keywords: MDSCs; autophagy; cancer; fatty acid oxidation; immune suppression; metabolism; oxidative phosphorylation; prostaglandin E2; stress; β-adrenergic signaling.
Copyright © 2021 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- Abuatiq A, Brown R, Wolles B, and Randall R (2020). Perceptions of stress: patient and caregiver experiences with stressors during hospitalization. Clin. J. Oncol. Nurs 24, 51–57. - PubMed
-
- Biswas SK (2015). Metabolic reprogramming of immune cells in cancer progression. Immunity 43, 435–449. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
