

Molecular mechanism of modulating miR482b level in tomato with botrytis cinerea infection
- PMID: 34706648
- PMCID: PMC8555085
- DOI: 10.1186/s12870-021-03203-2
Molecular mechanism of modulating miR482b level in tomato with botrytis cinerea infection
Abstract
Background: Plant miRNAs are involved in the response to biotic and abiotic stresses by altering their expression levels, and they play an important role in the regulation of plant resistance to stress. However, the molecular mechanism that regulates the expression levels of miRNAs in plants with biotic and abiotic stress still needs to be explored. Previously, we found that the expression of the miR482 family was changed in tomato infected by Botrytis cinerea. In this study, we investigated and uncovered the mechanism underlying the response of miR482 to B. cinerea infection in tomato.
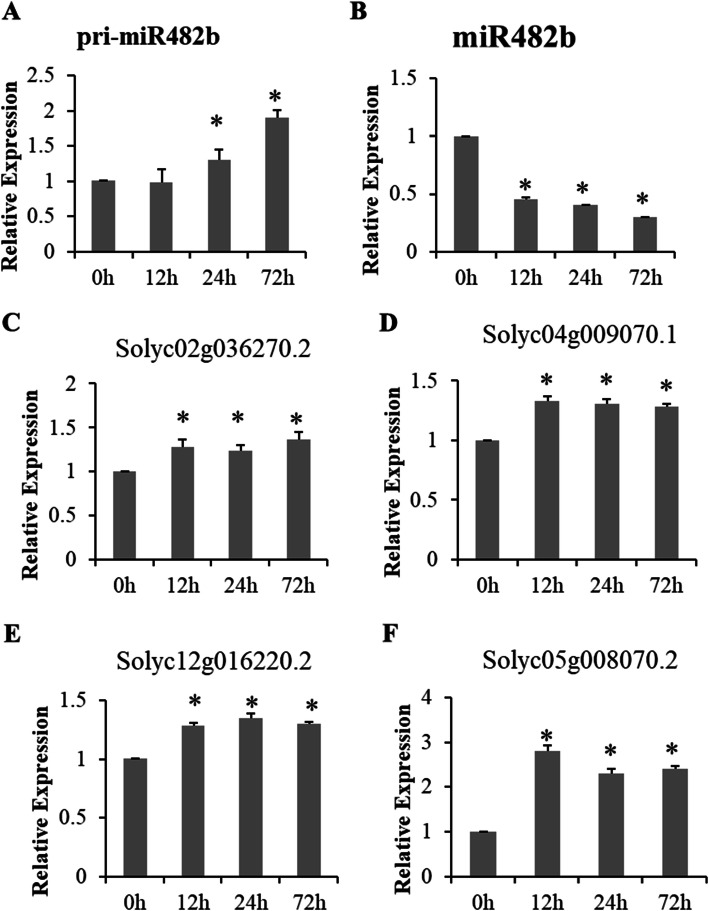
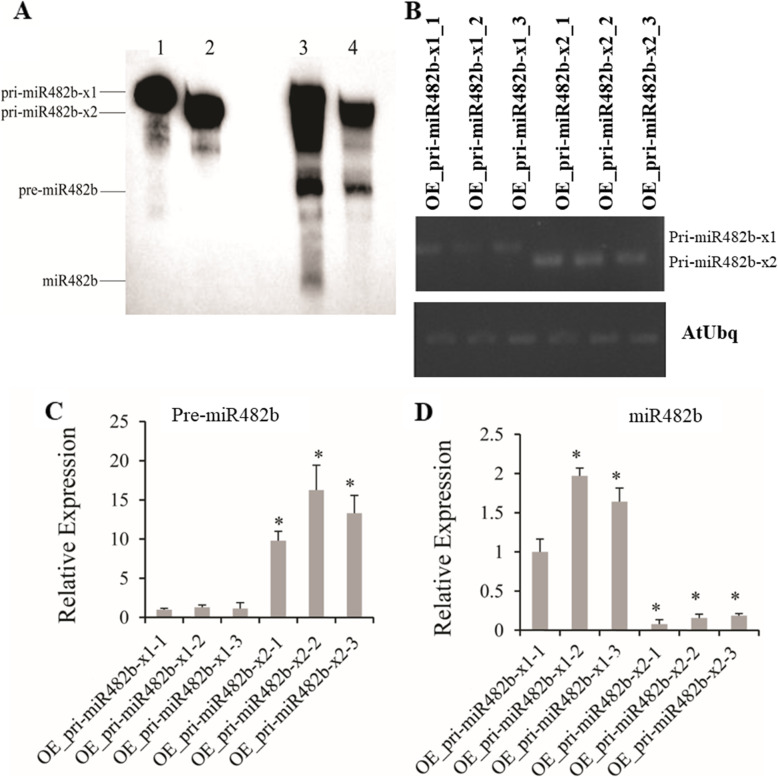
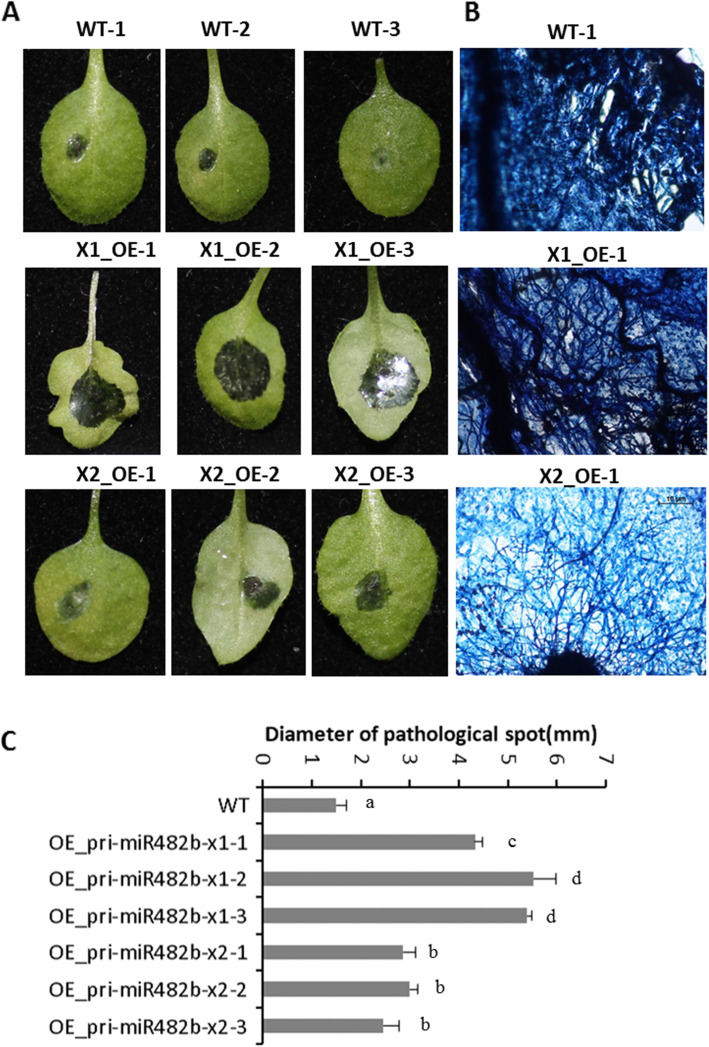
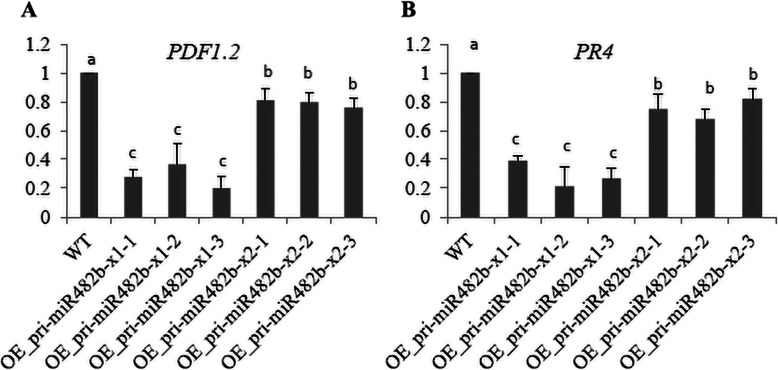
Results: First, RT-qPCR was employed to detect the expression patterns of miR482b in tomato infected by B. cinerea, and results showed that miR482b primary transcripts (pri-miR482b) were up-regulated in B. cinerea-infected leaves, but the mature miR482b was down-regulated. Subsequently, we used rapid amplification cDNA end method to amplify the full-length of pri-miR482b. Result showed that the pri-miR482b had two isoforms, with the longer one (consisting 300 bp) having an extra fragment of 53 bp in the 3'-end compared with the shorter one. In vitro Dicer assay indicated that the longer isoform pri-miR482b-x1 had higher efficiency in the post-transcriptional splicing of miRNA than the shorter isoform pri-miR482b-x2. In addition, the transcription level of mature miR482b was much higher in transgenic Arabidopsis overexpressing pri-miR482b-x1 than that in OE pri-miR482b-x2 Arabidopsis. These results confirmed that this extra 53 bp in pri-miR482b-x1 might play a key role in the miR482b biogenesis of post-transcription processing.
Conclusions: Extra 53 bp in pri-miR482b-x1 enhanced miR482b biogenesis, which elevated the transcription level of miR482b. This study clarified the response of miR482 to B. cinerea infection in tomato, thereby helping us further understand the molecular mechanisms that regulate the expression levels of other miRNAs.
Keywords: Botrytis cinerea; pathogen response; posttranscriptional processing; pri-miR482b isoforms; sly-miR482b.
© 2021. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures


References
MeSH terms
Substances
Supplementary concepts
Grants and funding
- LY21C140004 and LY18C150010/This study was supported by the Natural Science Foundation of Zhejiang Province, and the 521 Talent Foundation of Zhejiang Sci-Tech University.
- LY21C140004 and LY18C150010/This study was supported by the Natural Science Foundation of Zhejiang Province, and the 521 Talent Foundation of Zhejiang Sci-Tech University.
- LY21C140004 and LY18C150010/This study was supported by the Natural Science Foundation of Zhejiang Province, and the 521 Talent Foundation of Zhejiang Sci-Tech University.
- LY21C140004 and LY18C150010/This study was supported by the Natural Science Foundation of Zhejiang Province, and the 521 Talent Foundation of Zhejiang Sci-Tech University.
- LY21C140004 and LY18C150010/This study was supported by the Natural Science Foundation of Zhejiang Province, and the 521 Talent Foundation of Zhejiang Sci-Tech University.
LinkOut - more resources
Full Text Sources
