

On the role of dauer in the adaptation of nematodes to a parasitic lifestyle
- PMID: 34706780
- PMCID: PMC8555053
- DOI: 10.1186/s13071-021-04953-6
On the role of dauer in the adaptation of nematodes to a parasitic lifestyle
Abstract
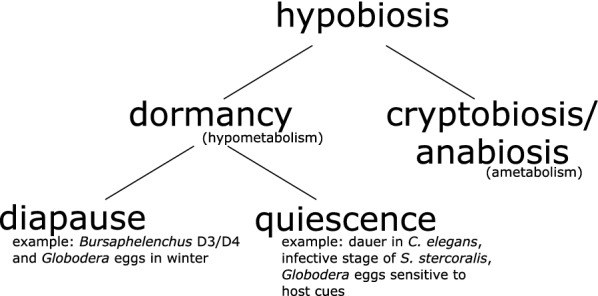
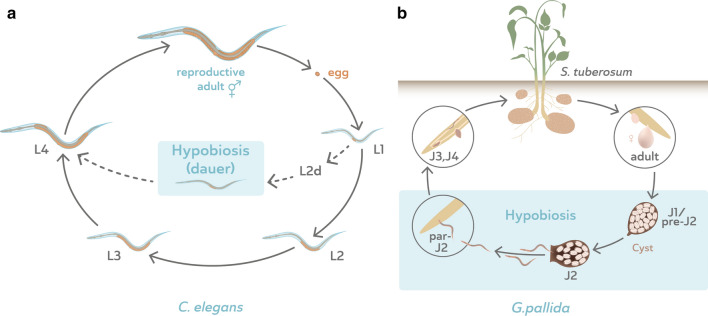
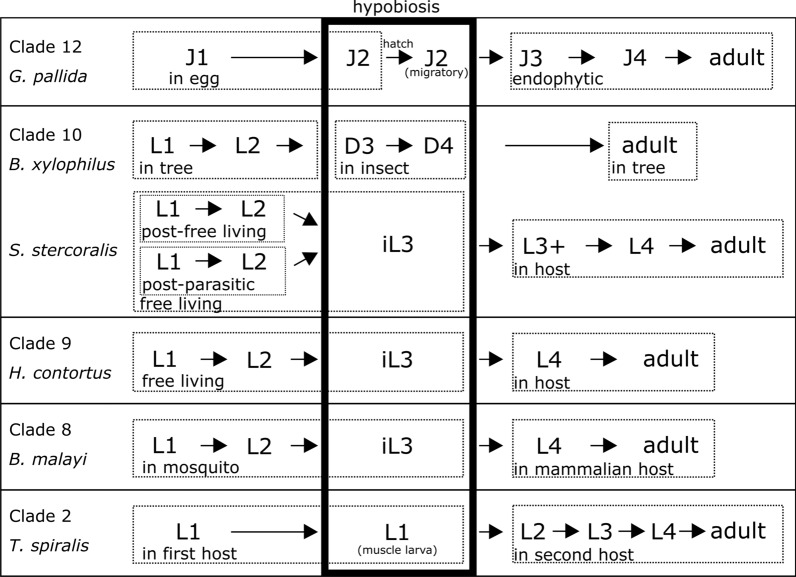
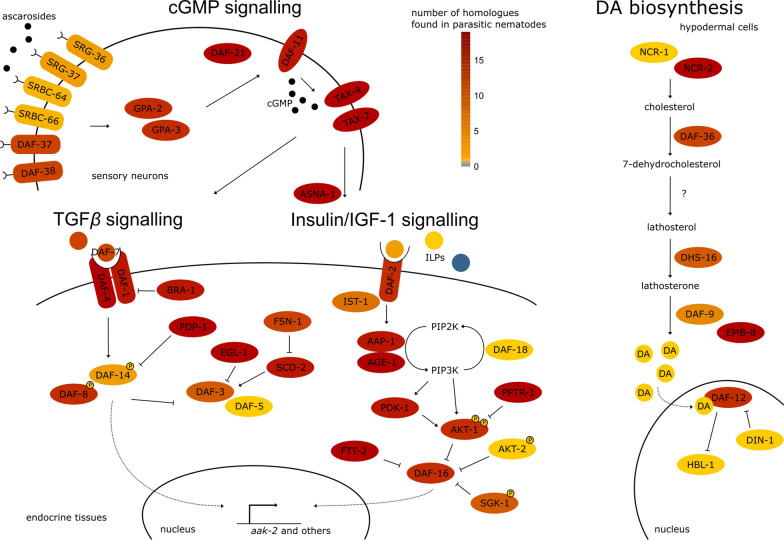
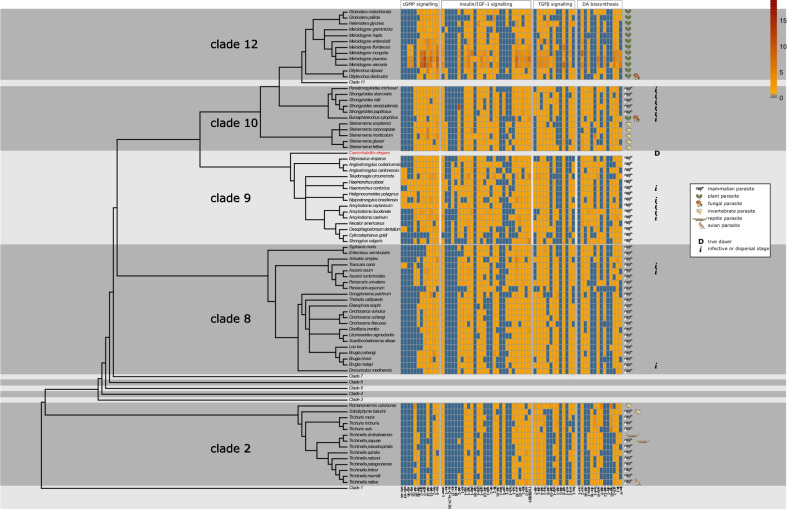
Nematodes are presumably the most abundant Metazoa on Earth, and can even be found in some of the most hostile environments of our planet. Various types of hypobiosis evolved to adapt their life cycles to such harsh environmental conditions. The five most distal major clades of the phylum Nematoda (Clades 8-12), formerly referred to as the Secernentea, contain many economically relevant parasitic nematodes. In this group, a special type of hypobiosis, dauer, has evolved. The dauer signalling pathway, which culminates in the biosynthesis of dafachronic acid (DA), is intensively studied in the free-living nematode Caenorhabditis elegans, and it has been hypothesized that the dauer stage may have been a prerequisite for the evolution of a wide range of parasitic lifestyles among other nematode species. Biosynthesis of DA is not specific for hypobiosis, but if it results in exit of the hypobiotic state, it is one of the main criteria to define certain behaviour as dauer. Within Clades 9 and 10, the involvement of DA has been validated experimentally, and dauer is therefore generally accepted to occur in those clades. However, for other clades, such as Clade 12, this has hardly been explored. In this review, we provide clarity on the nomenclature associated with hypobiosis and dauer across different nematological subfields. We discuss evidence for dauer-like stages in Clades 8 to 12 and support this with a meta-analysis of available genomic data. Furthermore, we discuss indications for a simplified dauer signalling pathway in parasitic nematodes. Finally, we zoom in on the host cues that induce exit from the hypobiotic stage and introduce two hypotheses on how these signals might feed into the dauer signalling pathway for plant-parasitic nematodes. With this work, we contribute to the deeper understanding of the molecular mechanisms underlying hypobiosis in parasitic nematodes. Based on this, novel strategies for the control of parasitic nematodes can be developed.
Keywords: Clade 12; Dauer; Globodera; Parasitic nematodes; Quiescence.
© 2021. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
