

The genomic origins of the Bronze Age Tarim Basin mummies
- PMID: 34707286
- PMCID: PMC8580821
- DOI: 10.1038/s41586-021-04052-7
The genomic origins of the Bronze Age Tarim Basin mummies
Abstract
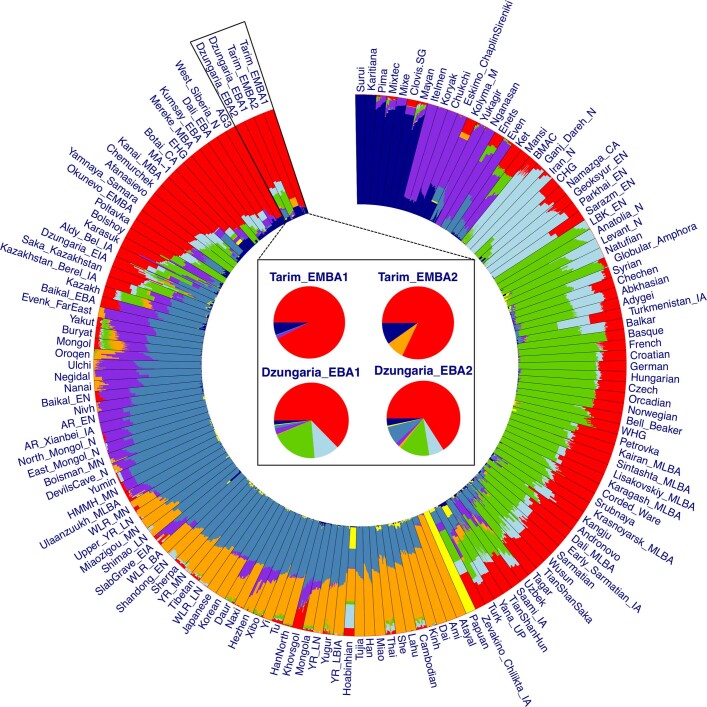
The identity of the earliest inhabitants of Xinjiang, in the heart of Inner Asia, and the languages that they spoke have long been debated and remain contentious1. Here we present genomic data from 5 individuals dating to around 3000-2800 BC from the Dzungarian Basin and 13 individuals dating to around 2100-1700 BC from the Tarim Basin, representing the earliest yet discovered human remains from North and South Xinjiang, respectively. We find that the Early Bronze Age Dzungarian individuals exhibit a predominantly Afanasievo ancestry with an additional local contribution, and the Early-Middle Bronze Age Tarim individuals contain only a local ancestry. The Tarim individuals from the site of Xiaohe further exhibit strong evidence of milk proteins in their dental calculus, indicating a reliance on dairy pastoralism at the site since its founding. Our results do not support previous hypotheses for the origin of the Tarim mummies, who were argued to be Proto-Tocharian-speaking pastoralists descended from the Afanasievo1,2 or to have originated among the Bactria-Margiana Archaeological Complex3 or Inner Asian Mountain Corridor cultures4. Instead, although Tocharian may have been plausibly introduced to the Dzungarian Basin by Afanasievo migrants during the Early Bronze Age, we find that the earliest Tarim Basin cultures appear to have arisen from a genetically isolated local population that adopted neighbouring pastoralist and agriculturalist practices, which allowed them to settle and thrive along the shifting riverine oases of the Taklamakan Desert.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures







Comment in
-
The unexpected ancestry of Inner Asian mummies.Nature. 2021 Nov;599(7884):204-206. doi: 10.1038/d41586-021-02872-1. Nature. 2021. PMID: 34707262 No abstract available.
-
DNA reveals surprise ancestry of mysterious Chinese mummies.Nature. 2021 Nov;599(7883):19-20. doi: 10.1038/d41586-021-02948-y. Nature. 2021. PMID: 34707265 No abstract available.
References
-
- Peyrot, M. in Aspects of Globalisation: Mobility, Exchange and the Development of Multi-Cultural States 12–17 (2017).
-
- Damgaard P, et al. 137 ancient human genomes from across the Eurasian steppes. Nature. 2018;557:369–374. - PubMed
-
- Hemphill BE, Mallory JP. Horse-mounted invaders from the Russo-Kazakh steppe or agricultural colonists from western Central Asia? A craniometric investigation of the Bronze Age settlement of Xinjiang. Am. J. Phys. Anthropol. 2004;124:199–222. - PubMed
-
- Betts A, Jia P, Abuduresule I. A new hypothesis for early Bronze Age cultural diversity in Xinjiang, China. Archaeol. Res. Asia. 2019;17:204–213.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
