

Emerging Contributions of Solid-State NMR Spectroscopy to Chromatin Structural Biology
- PMID: 34708075
- PMCID: PMC8544521
- DOI: 10.3389/fmolb.2021.741581
Emerging Contributions of Solid-State NMR Spectroscopy to Chromatin Structural Biology
Abstract
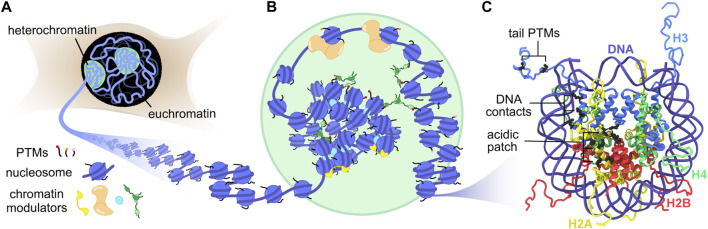
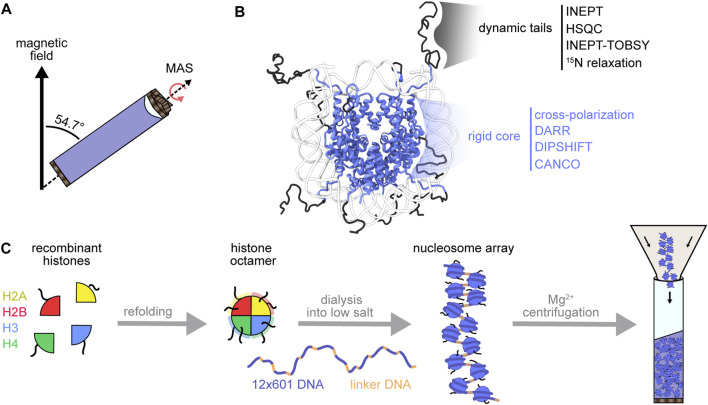
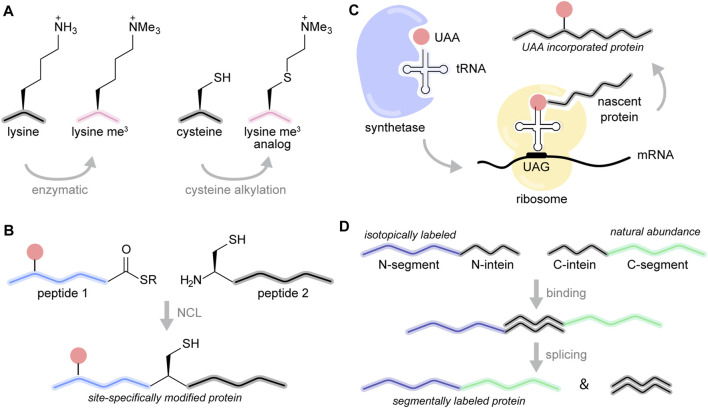
The eukaryotic genome is packaged into chromatin, a polymer of DNA and histone proteins that regulates gene expression and the spatial organization of nuclear content. The repetitive character of chromatin is diversified into rich layers of complexity that encompass DNA sequence, histone variants and post-translational modifications. Subtle molecular changes in these variables can often lead to global chromatin rearrangements that dictate entire gene programs with far reaching implications for development and disease. Decades of structural biology advances have revealed the complex relationship between chromatin structure, dynamics, interactions, and gene expression. Here, we focus on the emerging contributions of magic-angle spinning solid-state nuclear magnetic resonance spectroscopy (MAS NMR), a relative newcomer on the chromatin structural biology stage. Unique among structural biology techniques, MAS NMR is ideally suited to provide atomic level information regarding both the rigid and dynamic components of this complex and heterogenous biological polymer. In this review, we highlight the advantages MAS NMR can offer to chromatin structural biologists, discuss sample preparation strategies for structural analysis, summarize recent MAS NMR studies of chromatin structure and dynamics, and close by discussing how MAS NMR can be combined with state-of-the-art chemical biology tools to reconstitute and dissect complex chromatin environments.
Keywords: chemical biology; gene regulation; histone dynamics; magic angle spinning; nucleosome dynamics.
Copyright © 2021 Ackermann and Debelouchina.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources