

Presynaptic HCN channels constrain GABAergic synaptic transmission in pyramidal cells of the medial prefrontal cortex
- PMID: 34709375
- PMCID: PMC8966777
- DOI: 10.1242/bio.058840
Presynaptic HCN channels constrain GABAergic synaptic transmission in pyramidal cells of the medial prefrontal cortex
Abstract
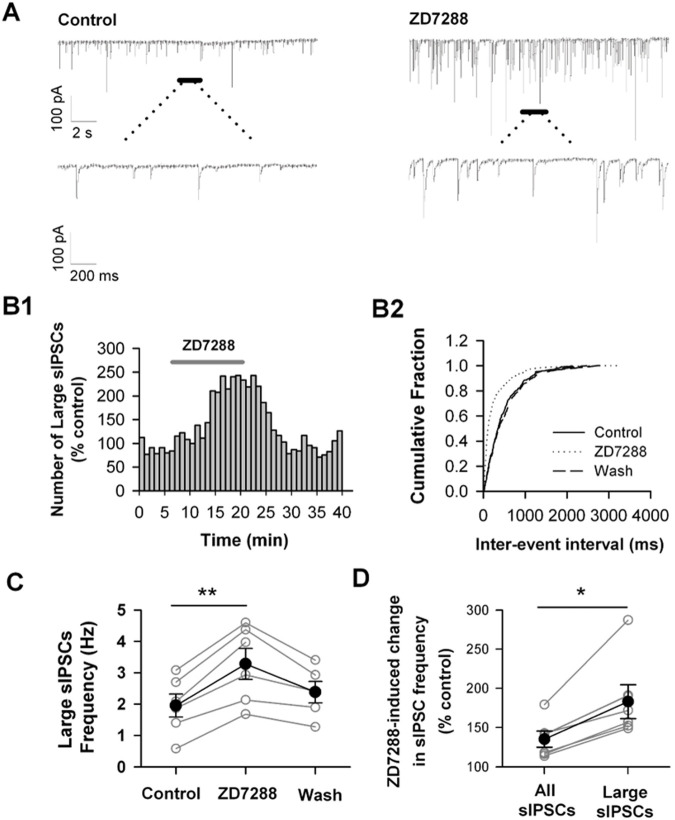
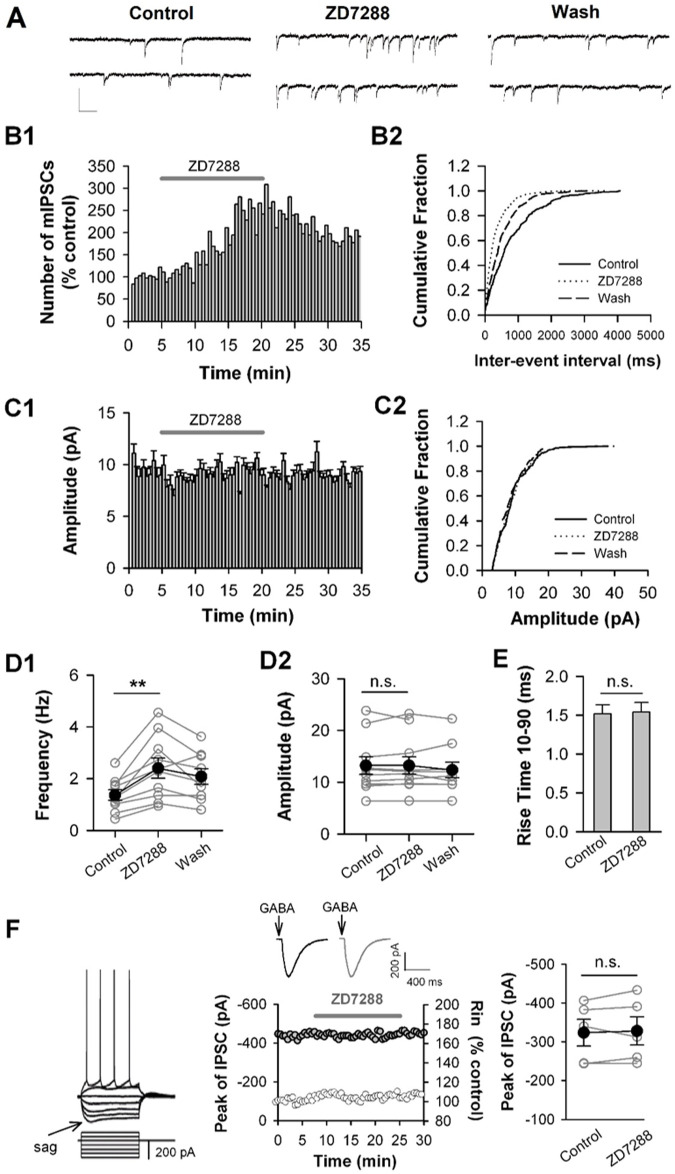
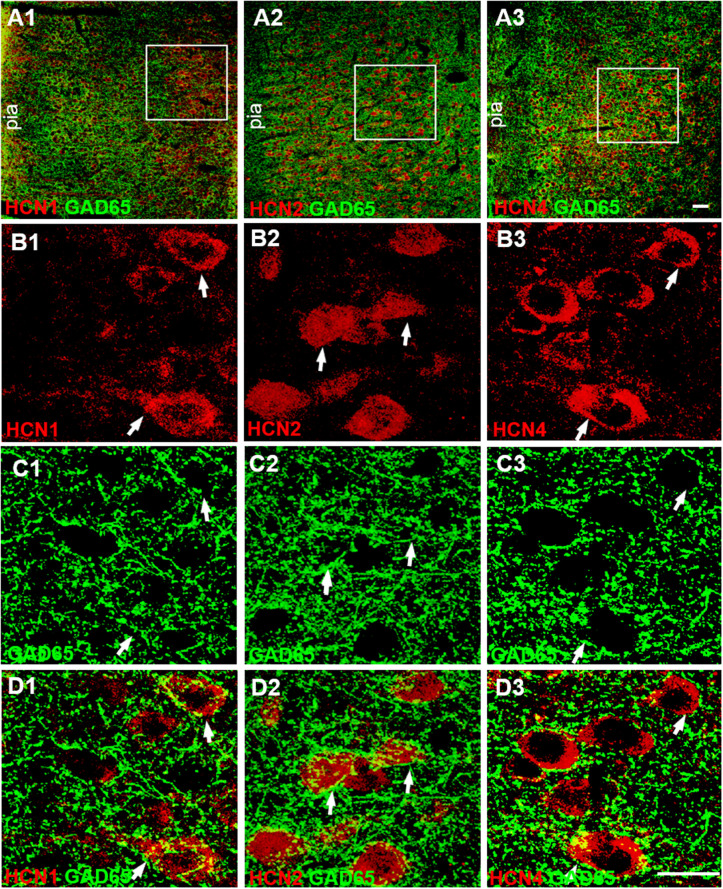
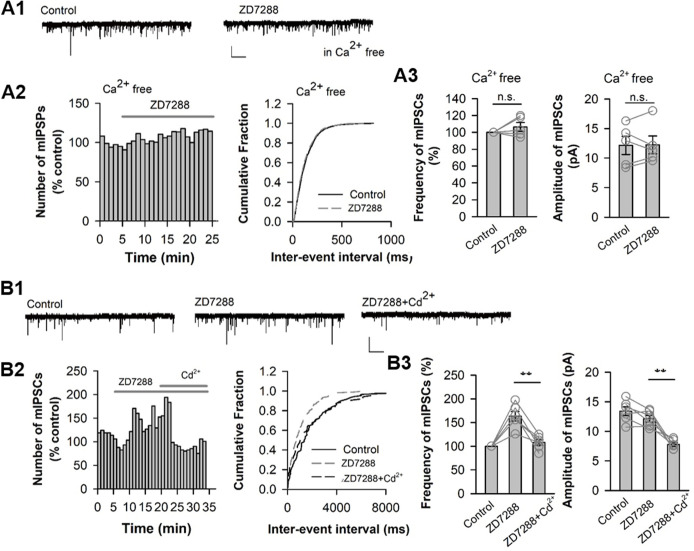
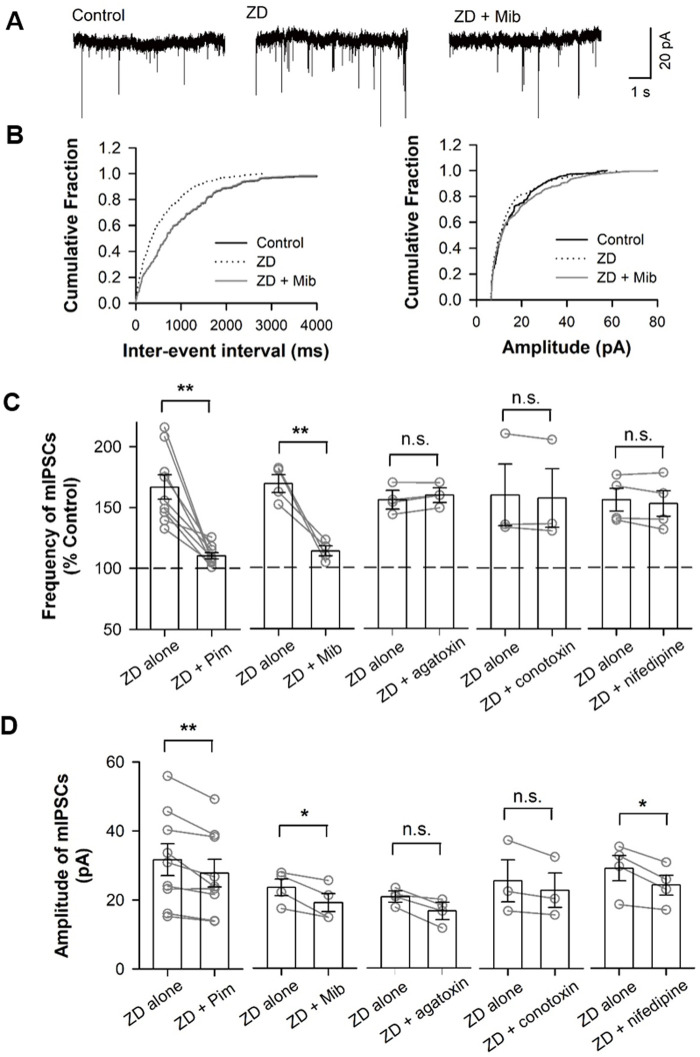
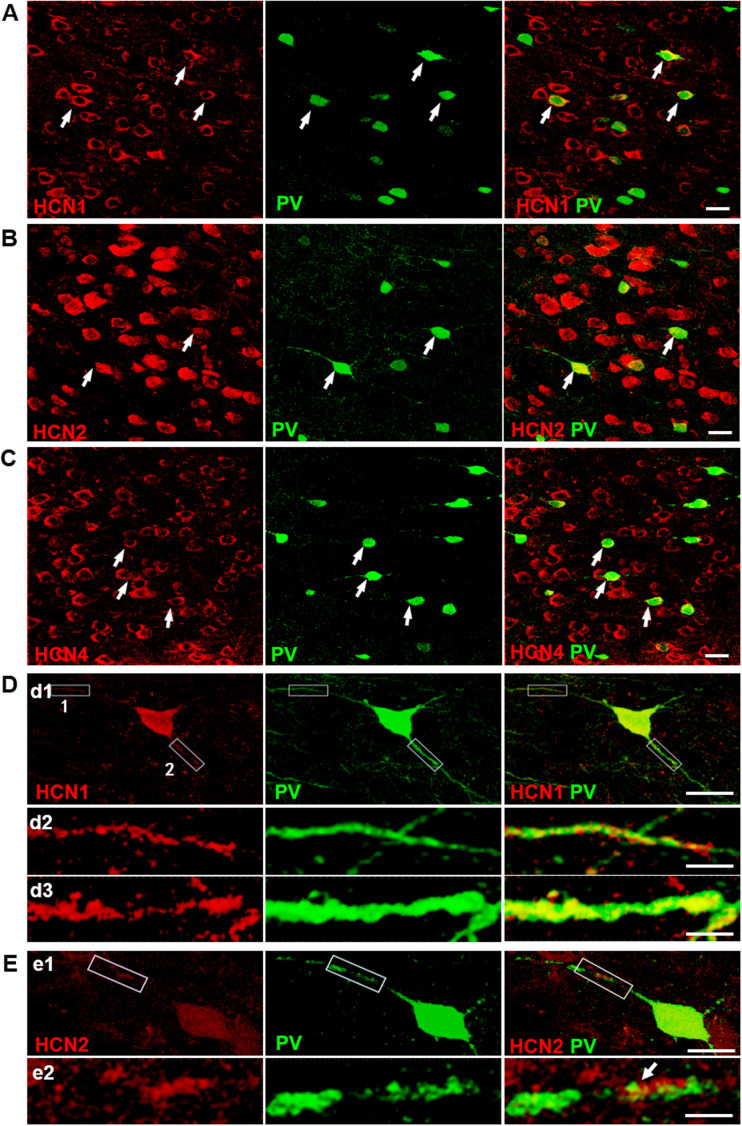
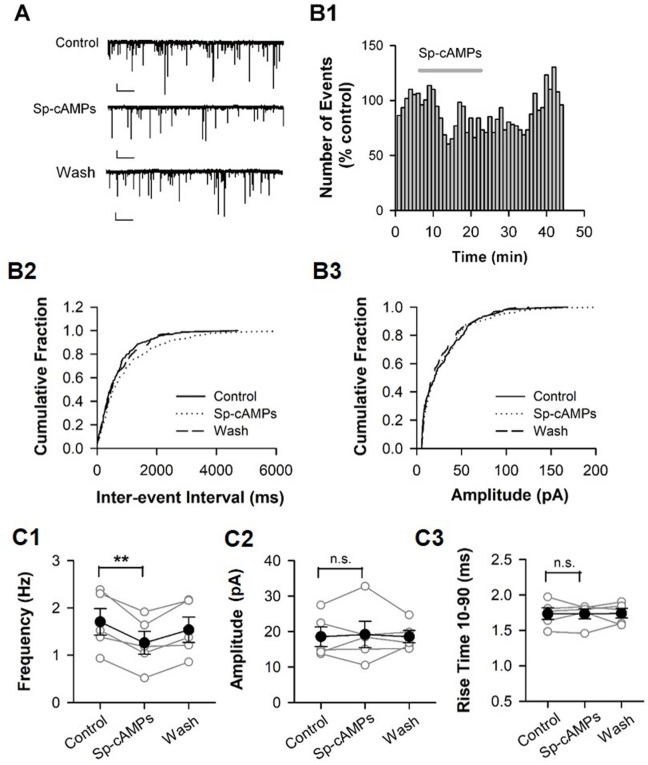
Hyperpolarization-activated cyclic nucleotide-gated (HCN) channels are widely expressed in neurons in the central nervous system. It has been documented that HCN channels regulate the intrinsic excitability of pyramidal cells in the medial prefrontal cortex (mPFC) of rodents. Here, we report that HCN channels limited GABAergic transmission onto pyramidal cells in rat mPFC. The pharmacological blockade of HCN channels resulted in a significant increase in the frequency of both spontaneous and miniature inhibitory postsynaptic currents (IPSCs) in mPFC pyramidal cells, whereas potentiation of HCN channels reversely decreases the frequency of mIPSCs. Furthermore, such facilitation effect on mIPSC frequency required presynaptic Ca2+ influx. Immunofluorescence staining showed that HCN channels expressed in presynaptic GABAergic terminals, as well as in both soma and neurite of parvalbumin-expressing (PV-expressing) basket cells in mPFC. The present results indicate that HCN channels in GABAergic interneurons, most likely PV-expressing basket cells, constrain inhibitory control over layer 5-6 pyramidal cells by restricting presynaptic Ca2+ entry.
Keywords: GABAergic transmission; HCN channel; Rats; mPFC.
© 2022. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interests The authors declare no competing or financial interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
