

A cis-regulatory element promoting increased transcription at low temperature in cultured ectothermic Drosophila cells
- PMID: 34711176
- PMCID: PMC8555087
- DOI: 10.1186/s12864-021-08057-4
A cis-regulatory element promoting increased transcription at low temperature in cultured ectothermic Drosophila cells
Erratum in
-
Correction to: A cis-regulatory element promoting increased transcription at low temperature in cultured ectothermic Drosophila cells.BMC Genomics. 2022 Mar 28;23(1):241. doi: 10.1186/s12864-022-08473-0. BMC Genomics. 2022. PMID: 35346028 Free PMC article. No abstract available.
Abstract
Background: Temperature change affects the myriad of concurrent cellular processes in a non-uniform, disruptive manner. While endothermic organisms minimize the challenge of ambient temperature variation by keeping the core body temperature constant, cells of many ectothermic species maintain homeostatic function within a considerable temperature range. The cellular mechanisms enabling temperature acclimation in ectotherms are still poorly understood. At the transcriptional level, the heat shock response has been analyzed extensively. The opposite, the response to sub-optimal temperature, has received lesser attention in particular in animal species. The tissue specificity of transcriptional responses to cool temperature has not been addressed and it is not clear whether a prominent general response occurs. Cis-regulatory elements (CREs), which mediate increased transcription at cool temperature, and responsible transcription factors are largely unknown.
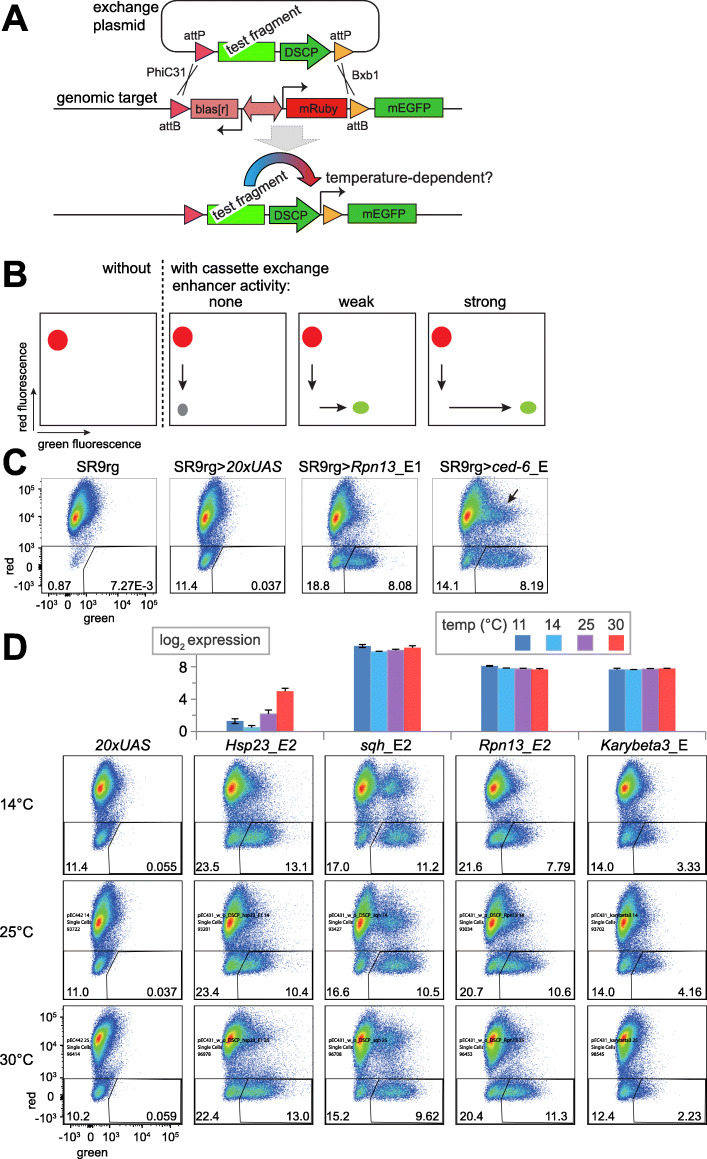
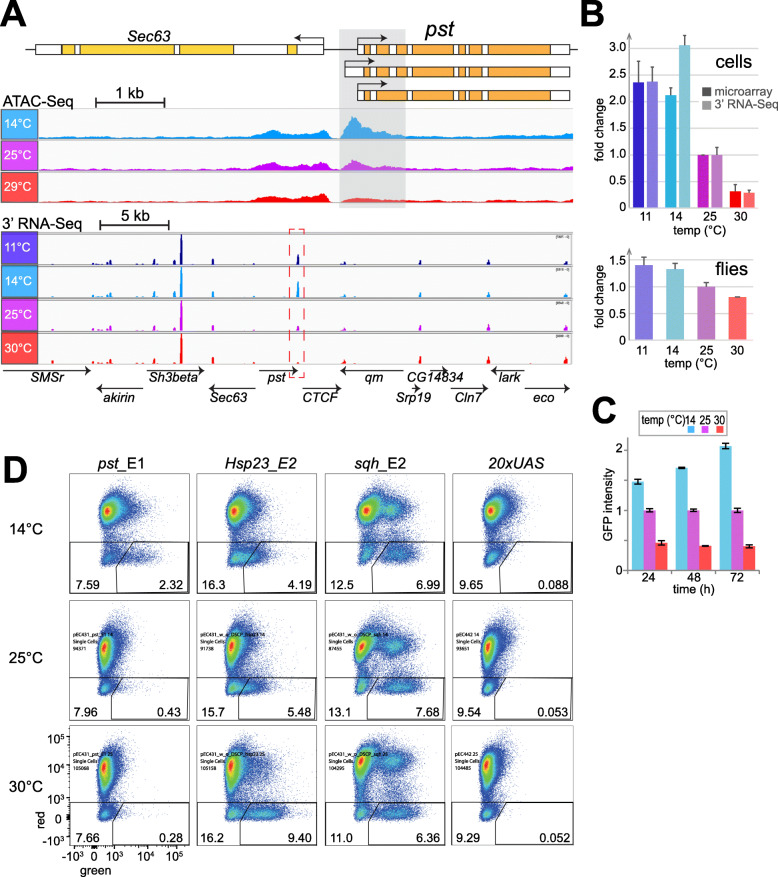
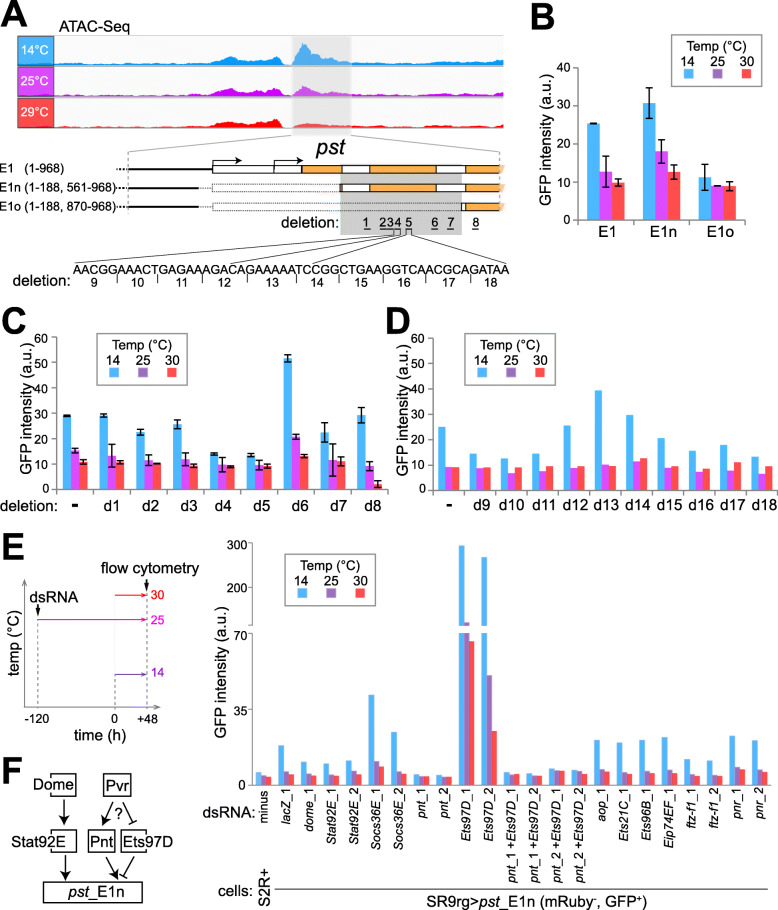
Results: The ectotherm Drosophila melanogaster with a presumed temperature optimum around 25 °C was used for transcriptomic analyses of effects of temperatures at the lower end of the readily tolerated range (14-29 °C). Comparative analyses with adult flies and cell culture lines indicated a striking degree of cell-type specificity in the transcriptional response to cool. To identify potential cis-regulatory elements (CREs) for transcriptional upregulation at cool temperature, we analyzed temperature effects on DNA accessibility in chromatin of S2R+ cells. Candidate cis-regulatory elements (CREs) were evaluated with a novel reporter assay for accurate assessment of their temperature-dependency. Robust transcriptional upregulation at low temperature could be demonstrated for a fragment from the pastrel gene, which expresses more transcript and protein at reduced temperatures. This CRE is controlled by the JAK/STAT signaling pathway and antagonizing activities of the transcription factors Pointed and Ets97D.
Conclusion: Beyond a rich data resource for future analyses of transcriptional control within the readily tolerated range of an ectothermic animal, a novel reporter assay permitting quantitative characterization of CRE temperature dependence was developed. Our identification and functional dissection of the pst_E1 enhancer demonstrate the utility of resources and assay. The functional characterization of this CoolUp enhancer provides initial mechanistic insights into transcriptional upregulation induced by a shift to temperatures at the lower end of the readily tolerated range.
Keywords: Cool temperature acclimation; Drosophila; Ectotherm; Ets97D; His2Av; NRF-2/GABP; S2R+ cells; Transcriptional control; cis-regulatory element (CRE); pastrel (pst).
© 2021. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
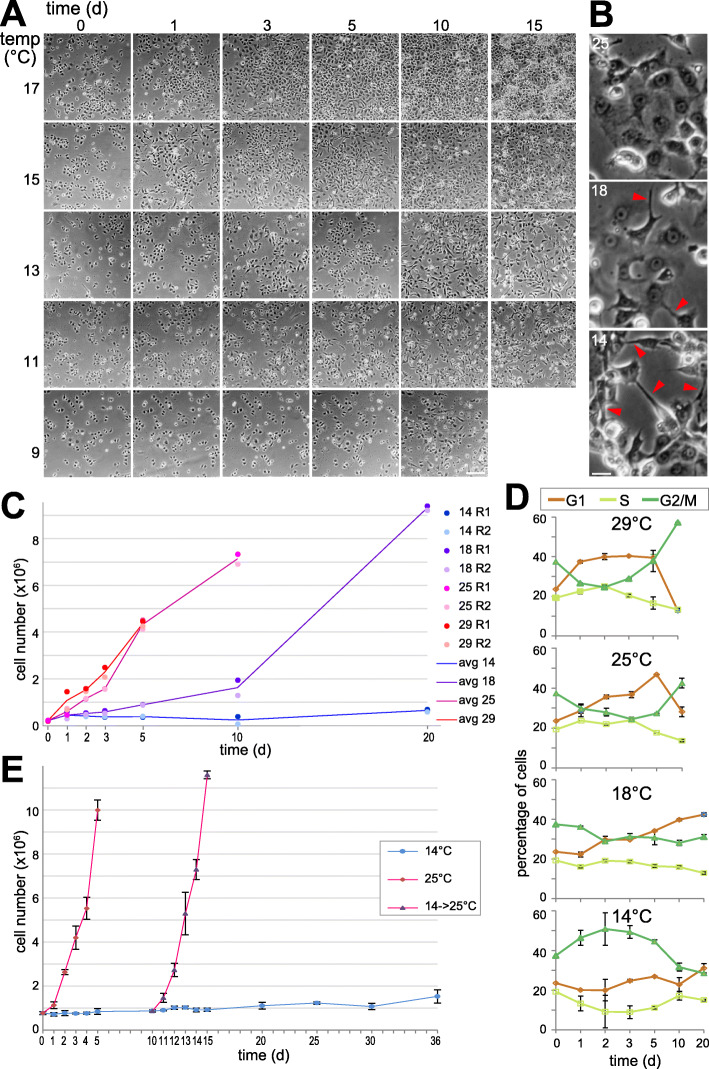
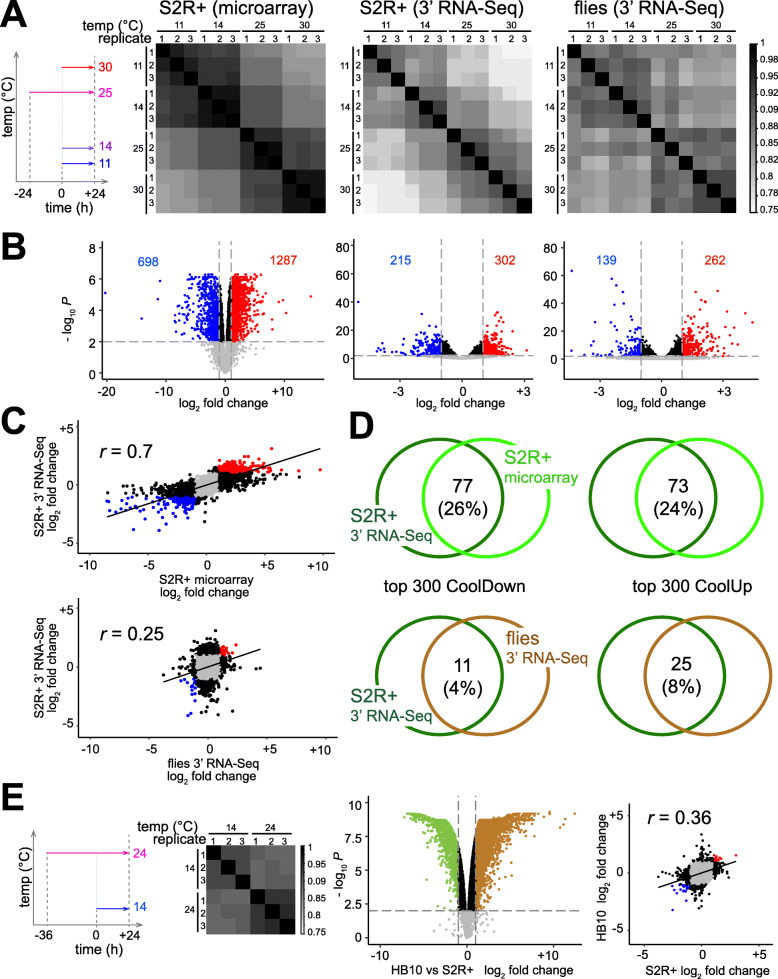
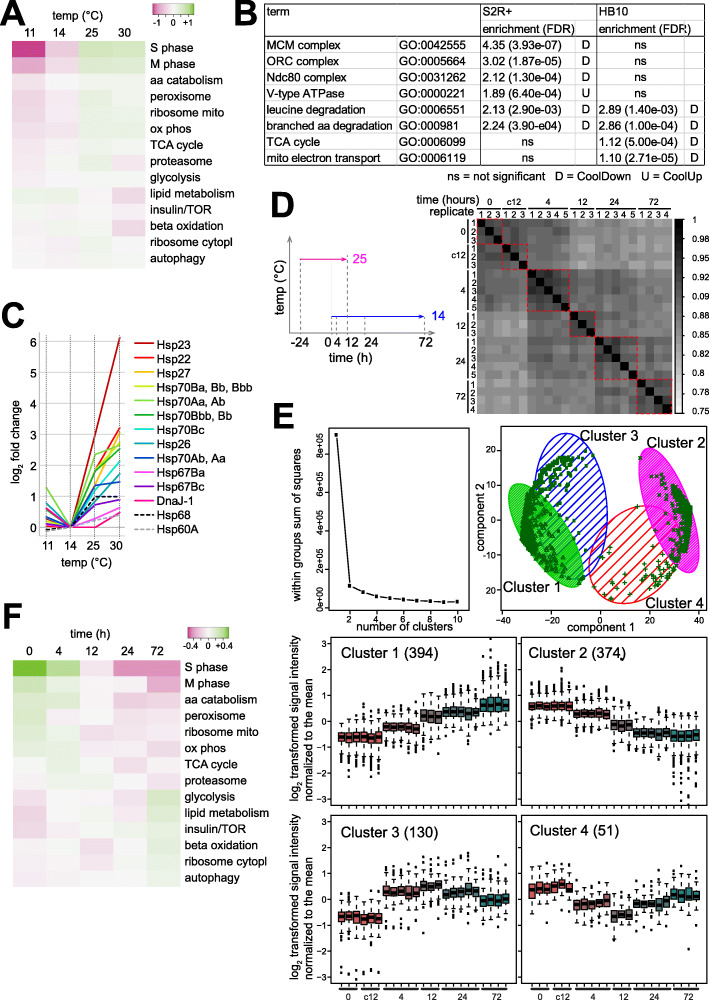
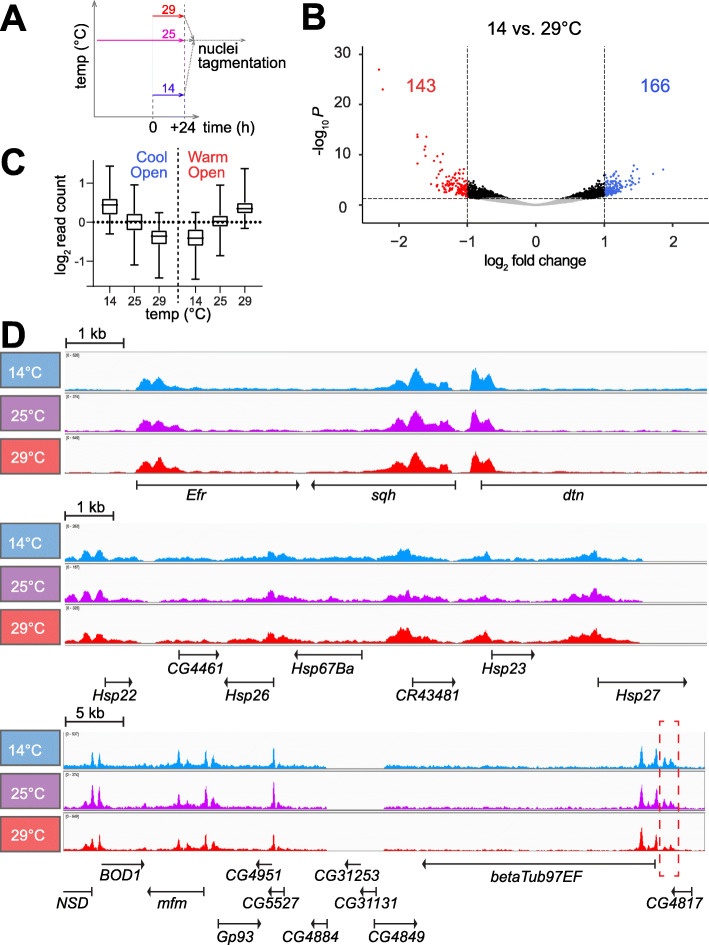
References
-
- Ritossa F. A new puffing pattern induced by temperature shock and DNP in Drosophila. Experientia. 1962;18(12):571–3. 10.1007/BF02172188.
-
- Liu AY, Bian H, Huang LE, Lee YK. Transient cold shock induces the heat shock response upon recovery at 37 degrees C in human cells. J Biol Chem. 1994;269(20):14768–75. 10.1016/S0021-9258(17)36691-7. - PubMed
-
- Colinet H, Hoffmann A. Gene and protein expression of Drosophila Starvin during cold stress and recovery from chill coma. Insect Biochem Mol Biol. 2010;40(5):425–8. 10.1016/j.ibmb.2010.03.002. - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
