

Metabolite discovery through global annotation of untargeted metabolomics data
- PMID: 34711973
- PMCID: PMC8733904
- DOI: 10.1038/s41592-021-01303-3
Metabolite discovery through global annotation of untargeted metabolomics data
Abstract
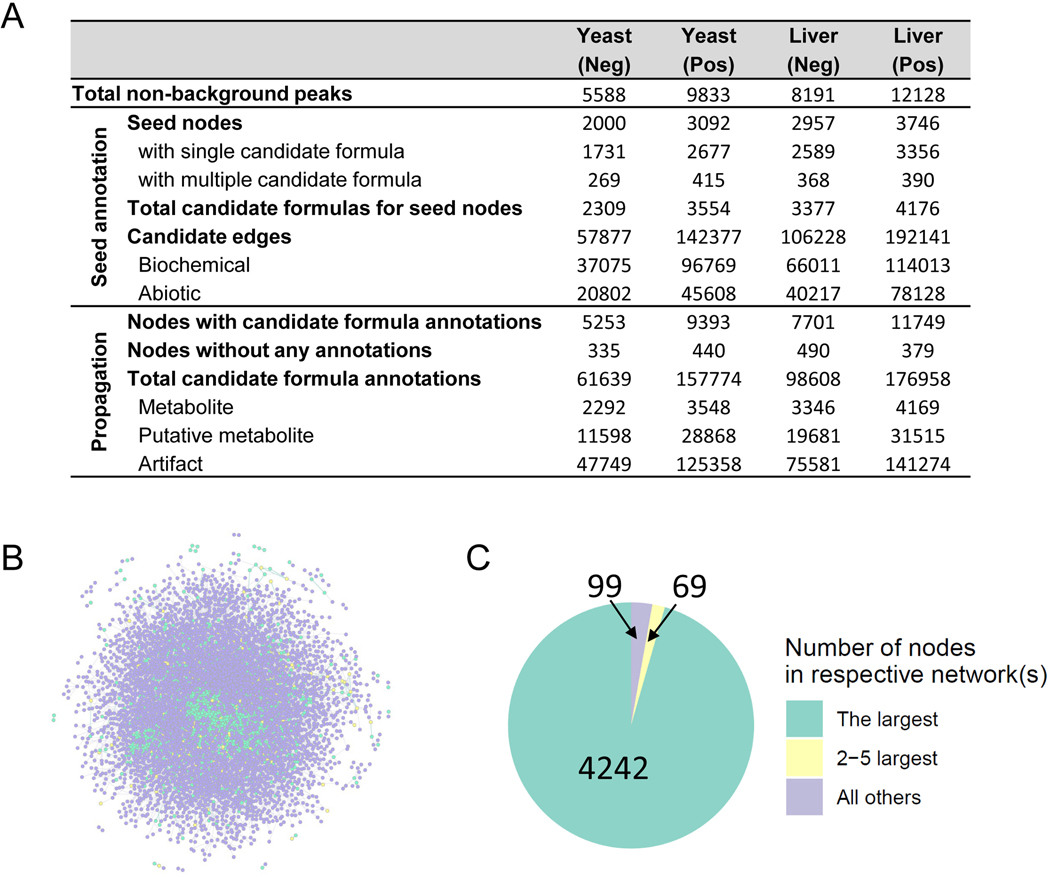
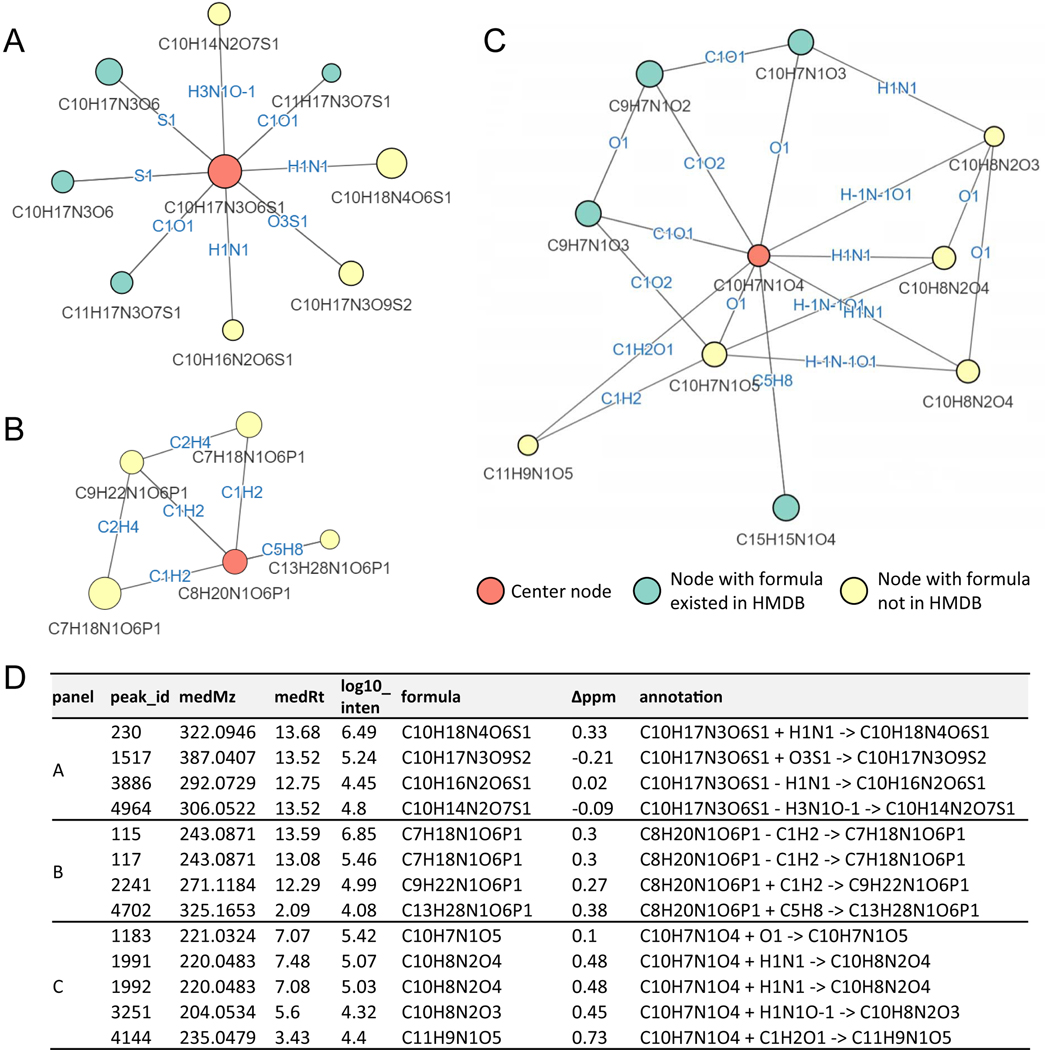
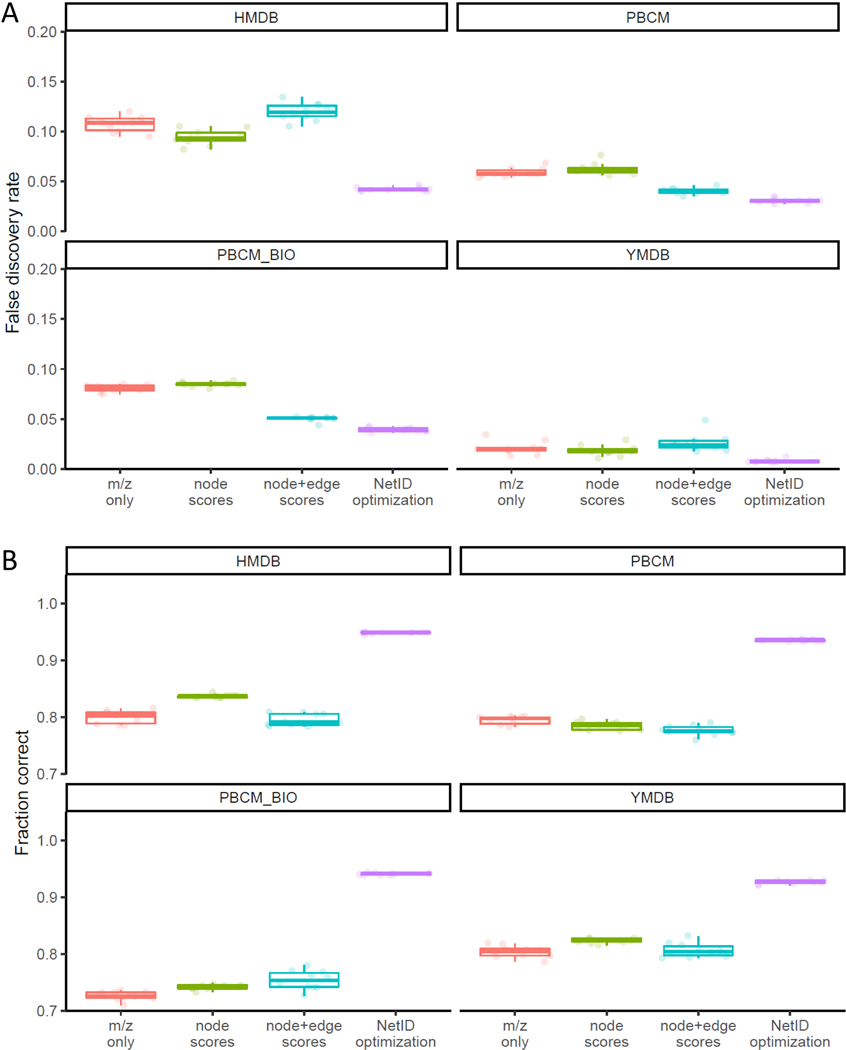
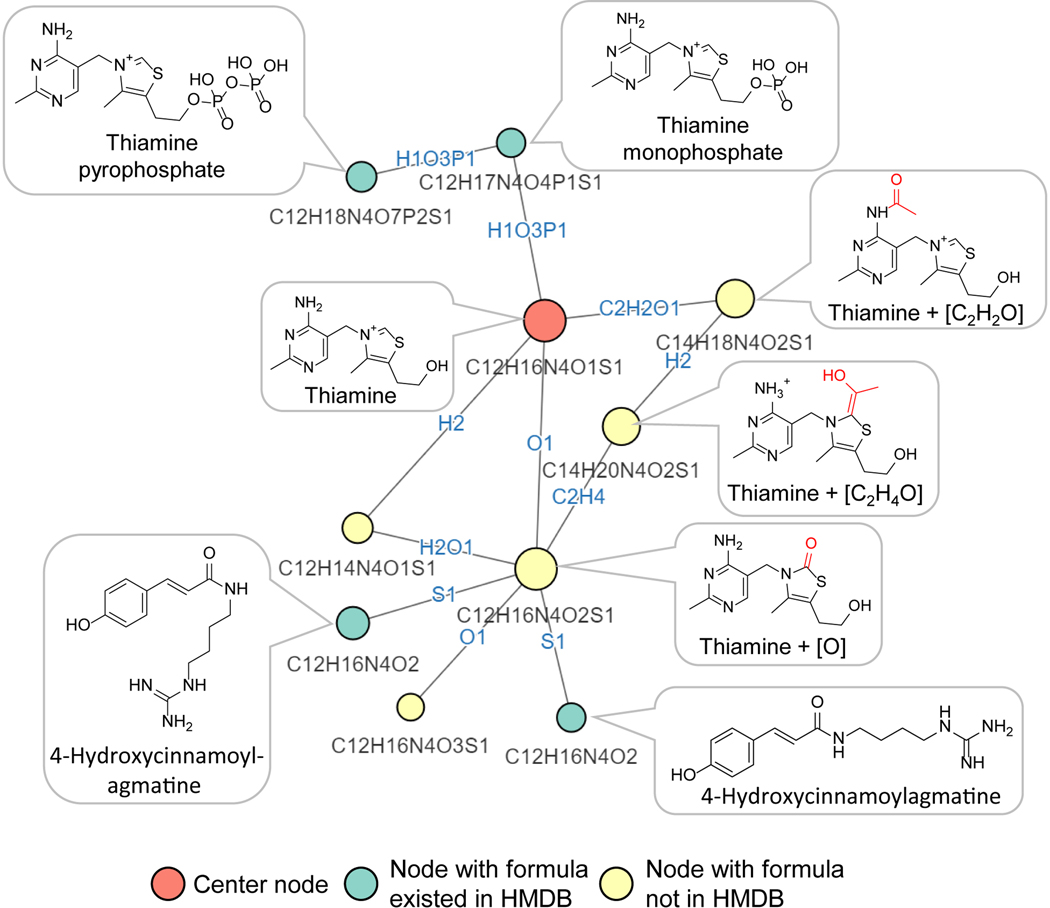
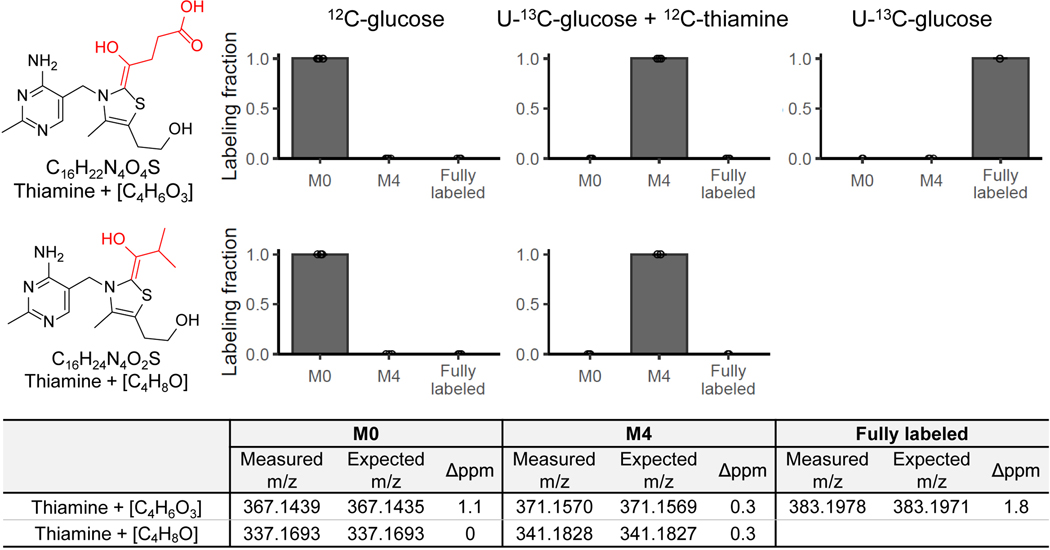
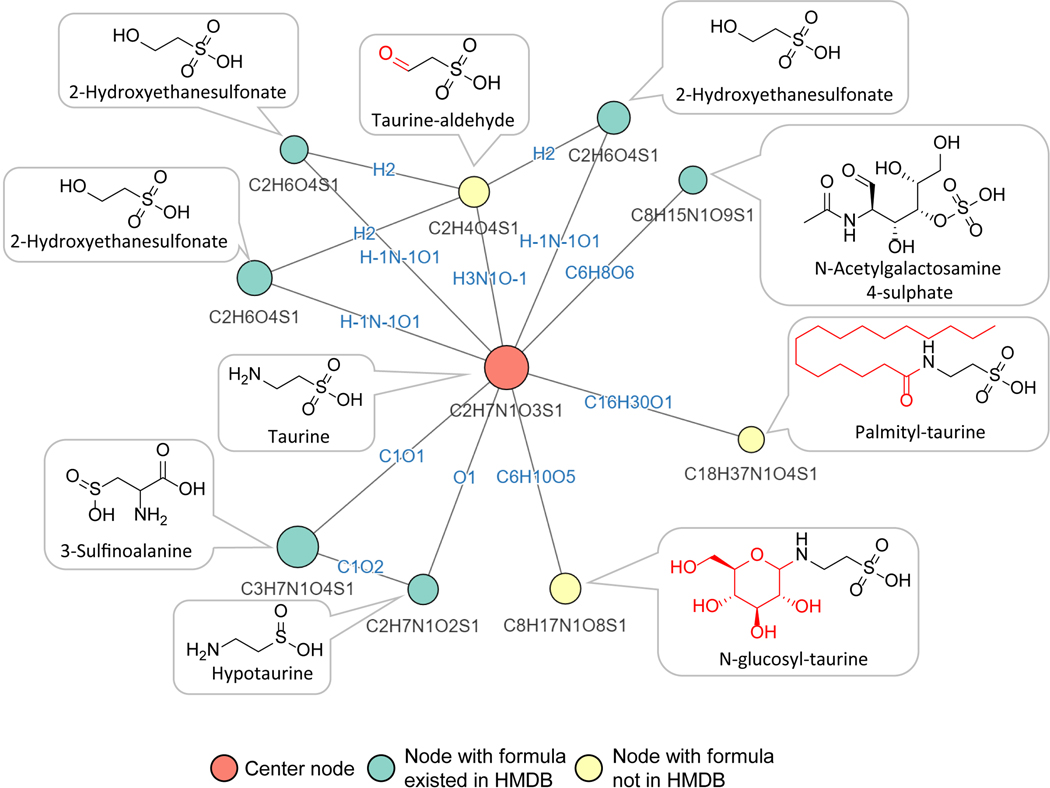
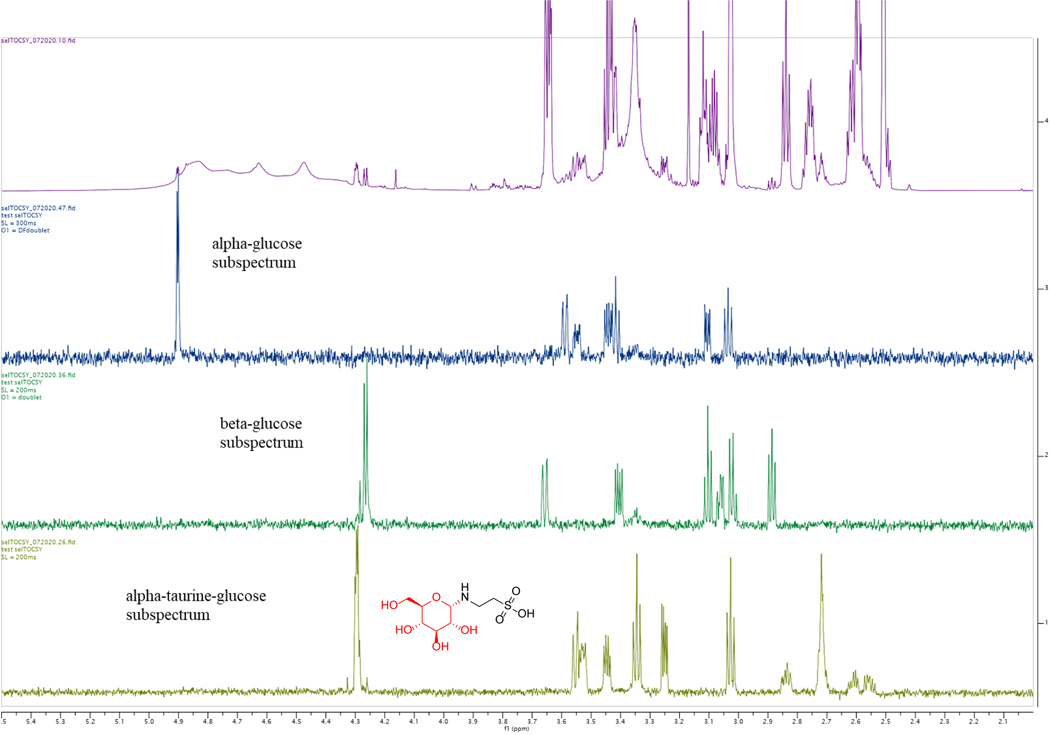
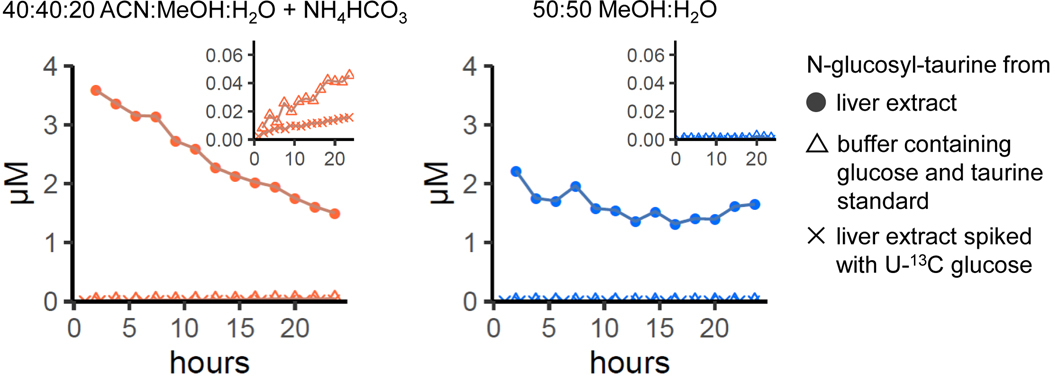
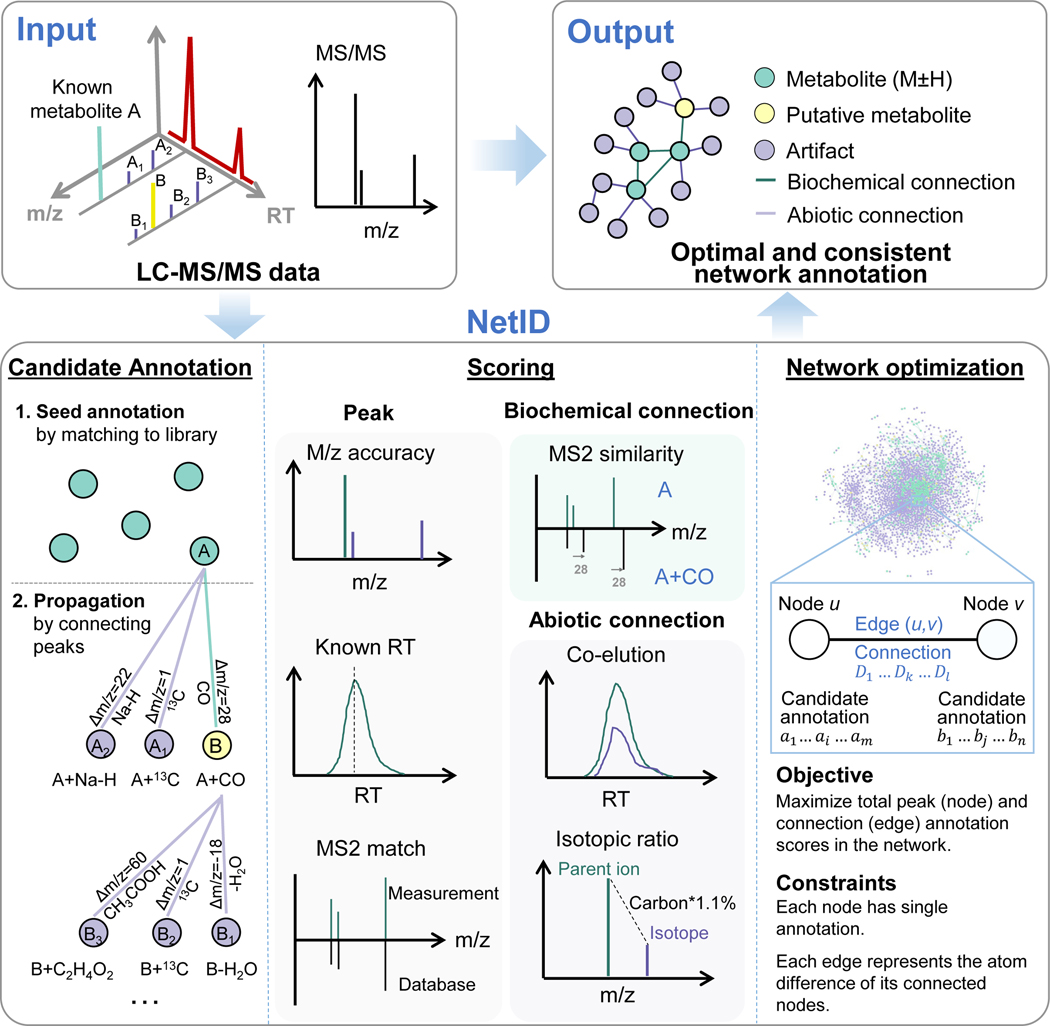
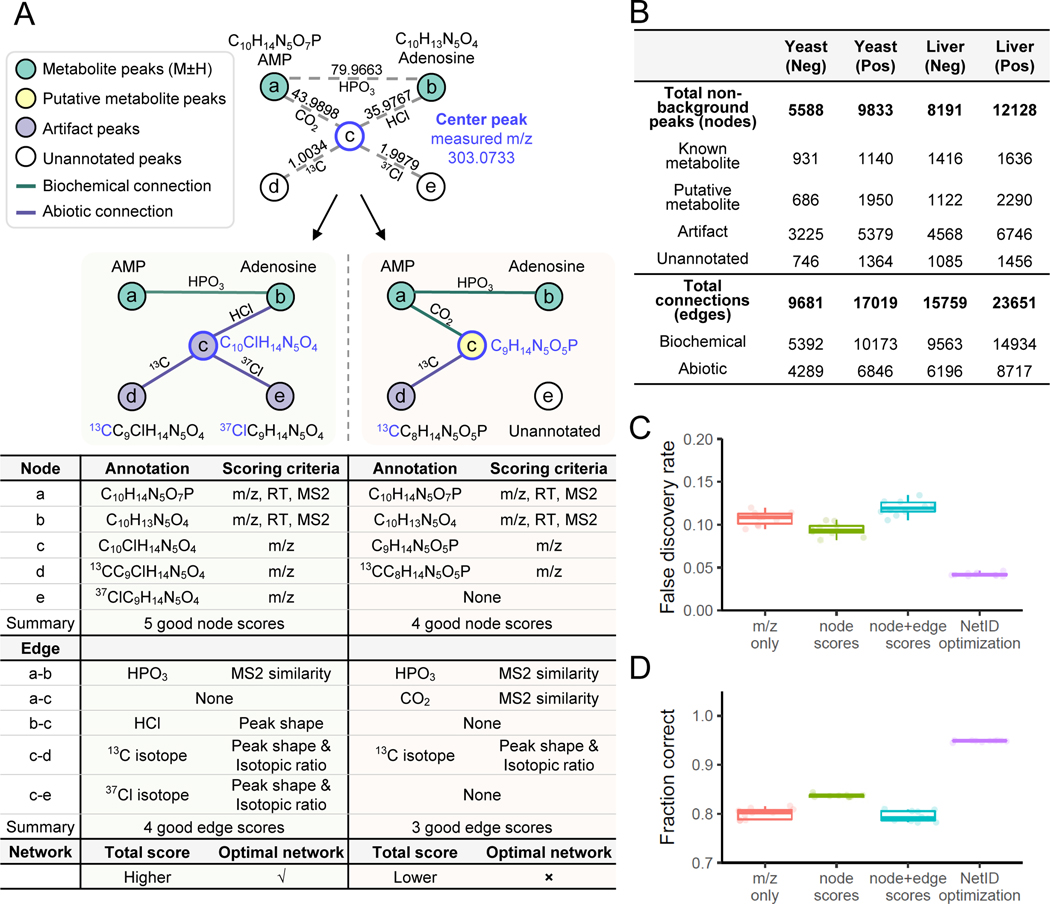
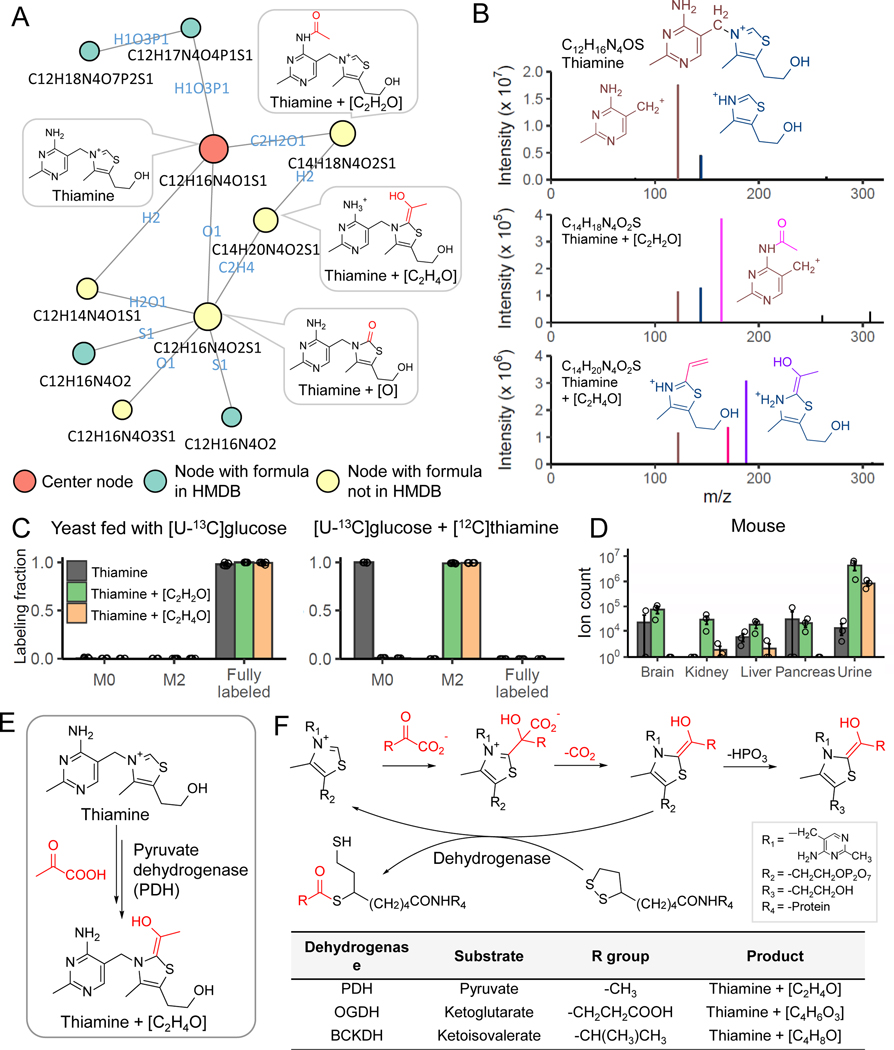
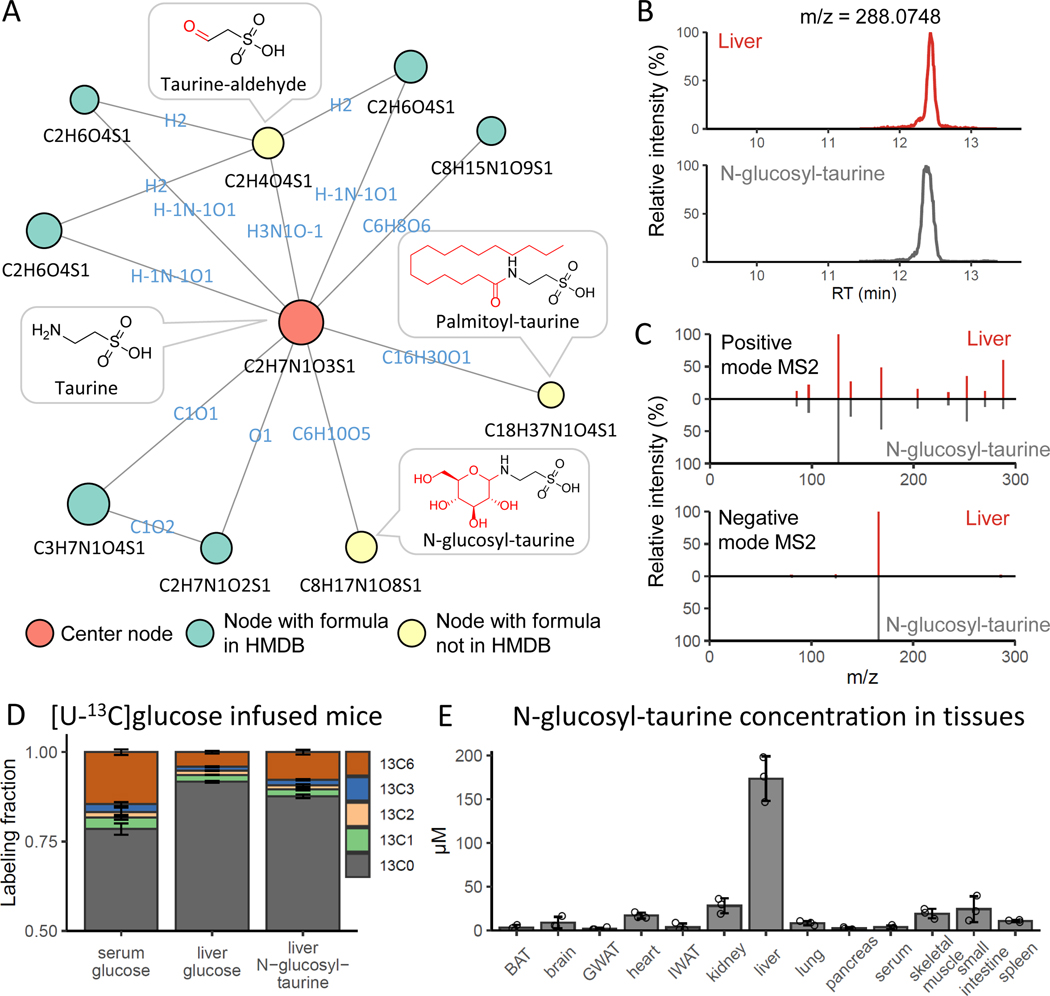
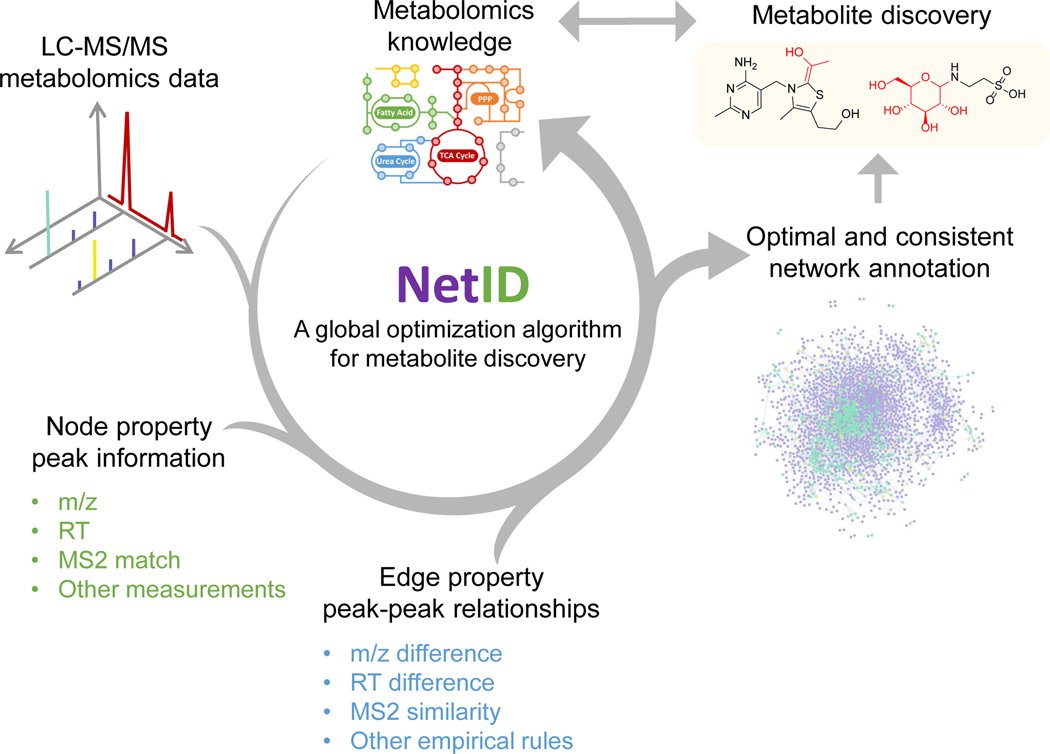
Liquid chromatography-high-resolution mass spectrometry (LC-MS)-based metabolomics aims to identify and quantify all metabolites, but most LC-MS peaks remain unidentified. Here we present a global network optimization approach, NetID, to annotate untargeted LC-MS metabolomics data. The approach aims to generate, for all experimentally observed ion peaks, annotations that match the measured masses, retention times and (when available) tandem mass spectrometry fragmentation patterns. Peaks are connected based on mass differences reflecting adduction, fragmentation, isotopes, or feasible biochemical transformations. Global optimization generates a single network linking most observed ion peaks, enhances peak assignment accuracy, and produces chemically informative peak-peak relationships, including for peaks lacking tandem mass spectrometry spectra. Applying this approach to yeast and mouse data, we identified five previously unrecognized metabolites (thiamine derivatives and N-glucosyl-taurine). Isotope tracer studies indicate active flux through these metabolites. Thus, NetID applies existing metabolomic knowledge and global optimization to substantially improve annotation coverage and accuracy in untargeted metabolomics datasets, facilitating metabolite discovery.
© 2021. The Author(s), under exclusive licence to Springer Nature America, Inc.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures
References
Methods Reference
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
