

DYW domain structures imply an unusual regulation principle in plant organellar RNA editing catalysis
- PMID: 34712911
- PMCID: PMC7611903
- DOI: 10.1038/s41929-021-00633-x
DYW domain structures imply an unusual regulation principle in plant organellar RNA editing catalysis
Abstract
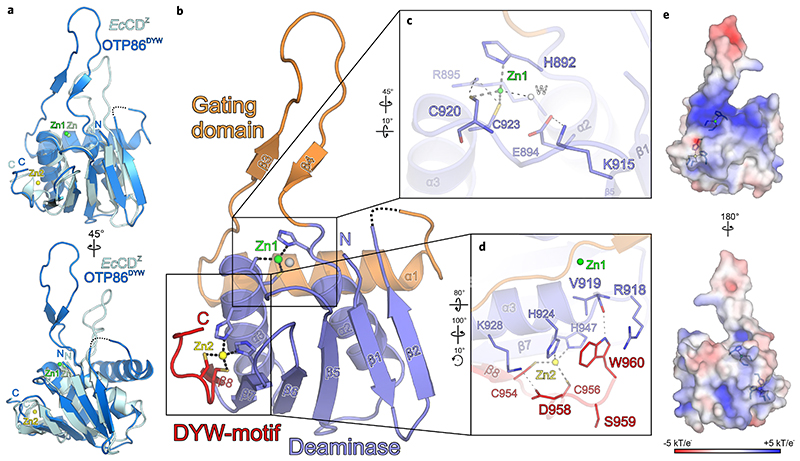
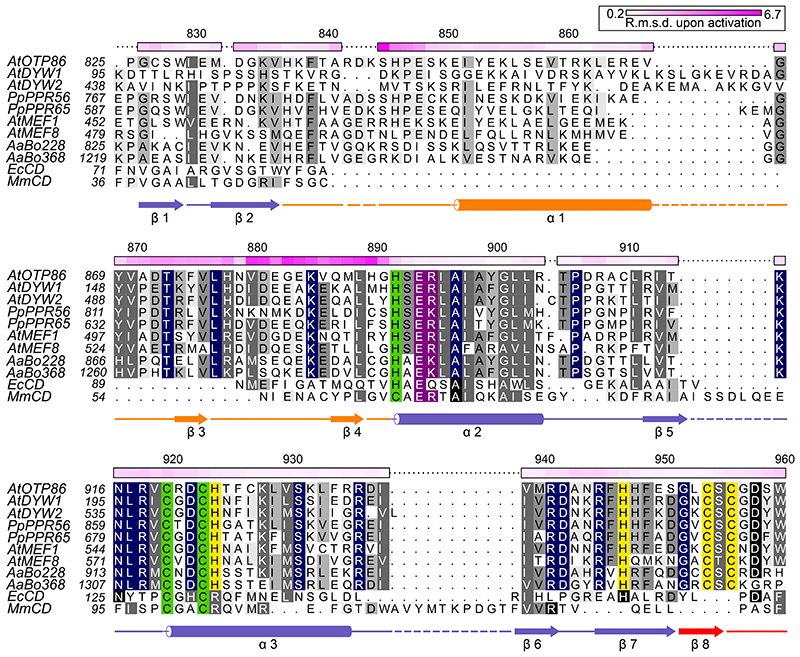
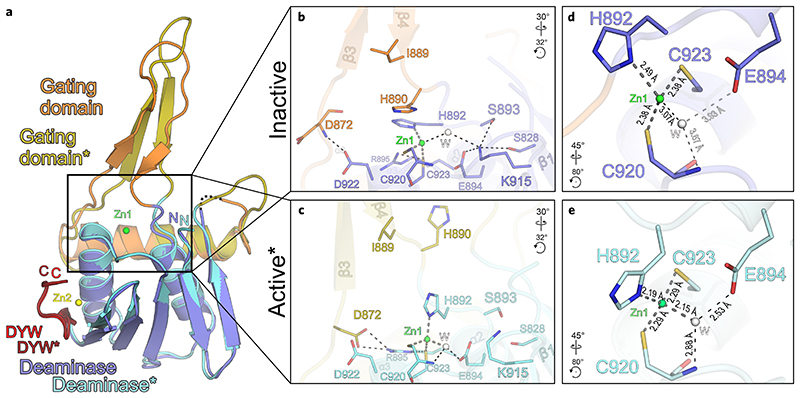
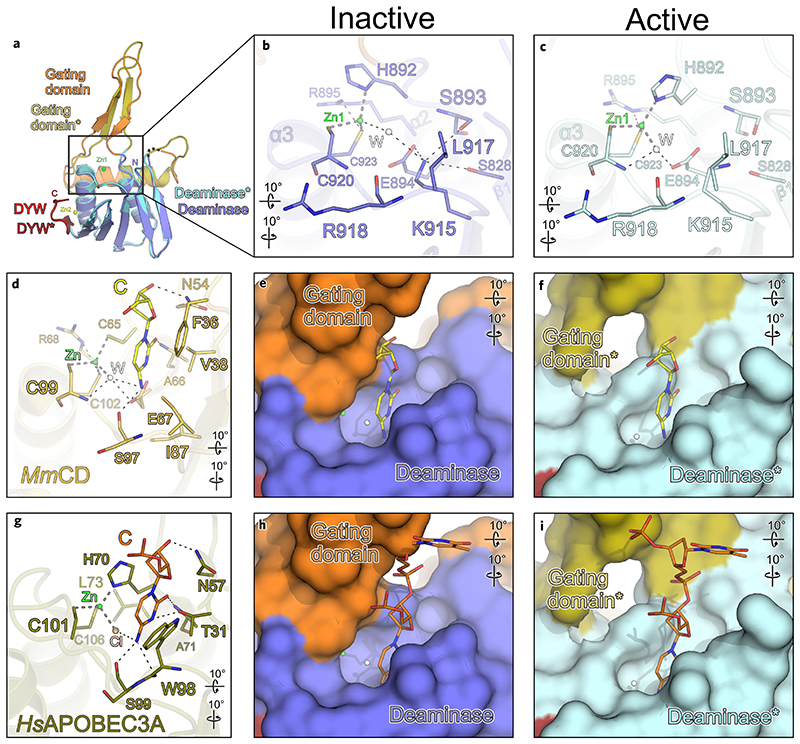
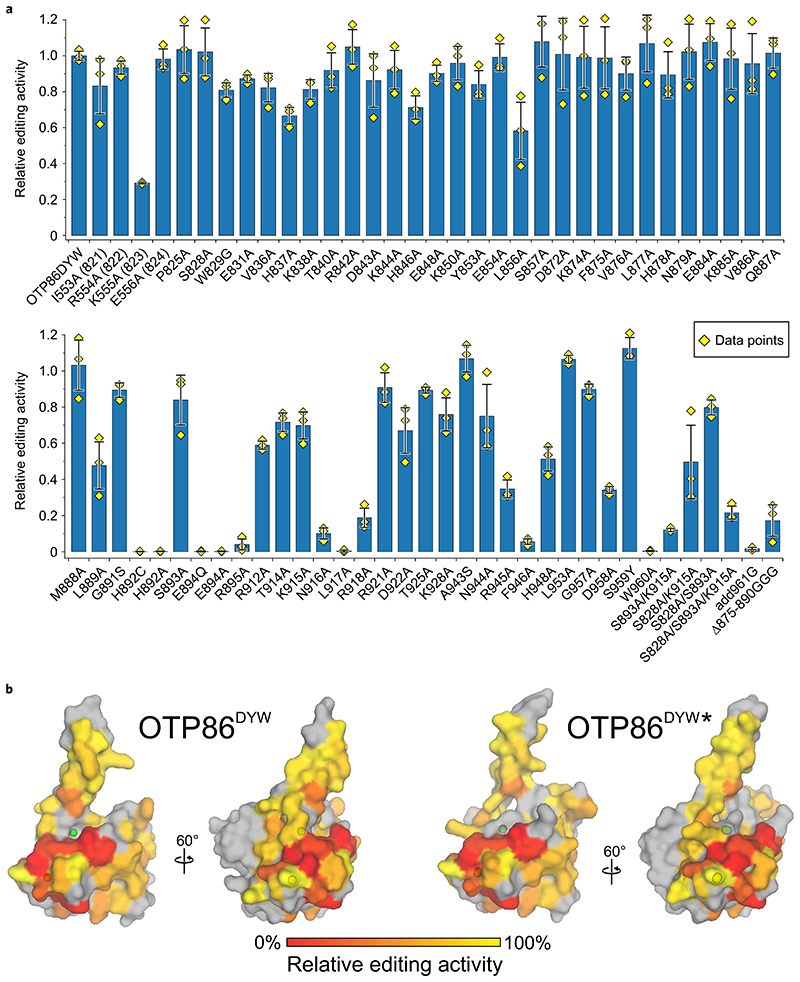
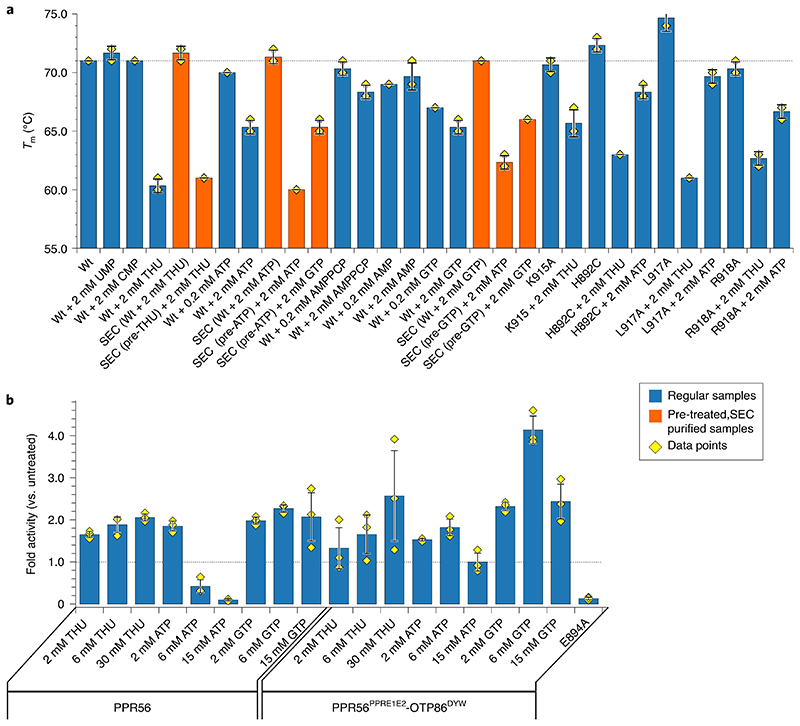
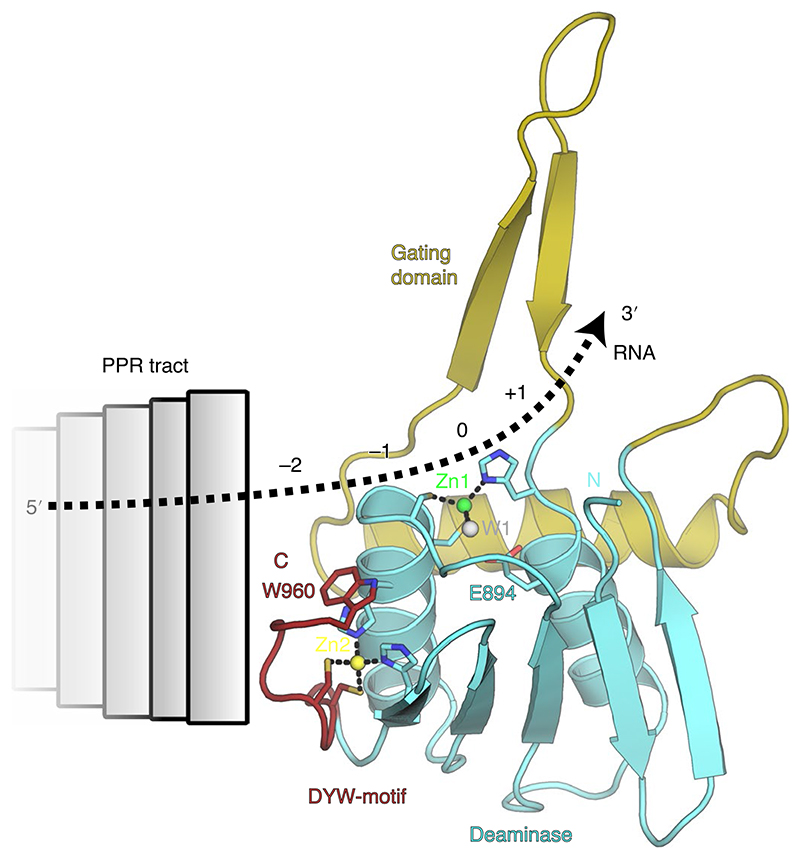
RNA editosomes selectively deaminate cytidines to uridines in plant organellar transcripts-mostly to restore protein functionality and consequently facilitate mitochondrial and chloroplast function. The RNA editosomal pentatricopeptide repeat proteins serve target RNA recognition, whereas the intensively studied DYW domain elicits catalysis. Here we present structures and functional data of a DYW domain in an inactive ground state and activated. DYW domains harbour a cytidine deaminase fold and a C-terminal DYW motif, with catalytic and structural zinc atoms, respectively. A conserved gating domain within the deaminase fold regulates the active site sterically and mechanistically in a process that we termed gated zinc shutter. Based on the structures, an autoinhibited ground state and its activation are cross-validated by RNA editing assays and differential scanning fluorimetry. We anticipate that, in vivo, the framework of an active plant RNA editosome triggers the release of DYW autoinhibition to ensure a controlled and coordinated cytidine deamination playing a key role in mitochondrial and chloroplast homeostasis.
Conflict of interest statement
Competing interests The authors declare no competing interests.
Figures
References
-
- Hiesel R, Wissinger B, Schuster W, Brennicke A. RNA editing in plant mitochondria. Science. 1989;246:1632–1634. - PubMed
-
- Covello PS, Gray MW. RNA editing in plant mitochondria. Nature. 1989;341:662–666. - PubMed
-
- Gualberto JM, Lamattina L, Bonnard G, Weil JH, Grienenberger JM. RNA editing in wheat mitochondria results in the conservation of protein sequences. Nature. 1989;341:660–662. - PubMed
-
- Hoch B, Maier RM, Appel K, Igloi GL, Kössel H. Editing of a chloroplast mRNA by creation of an initiation codon. Nature. 1991;353:178–180. - PubMed
-
- Takenaka M, Zehrmann A, Verbitskiy D, Härtel B, Brennicke A. RNA editing in plants and its evolution. Annu Rev Genet. 2013;47:335–352. - PubMed