

Kinetic proofreading of lipochitooligosaccharides determines signal activation of symbiotic plant receptors
- PMID: 34716271
- PMCID: PMC8612216
- DOI: 10.1073/pnas.2111031118
Kinetic proofreading of lipochitooligosaccharides determines signal activation of symbiotic plant receptors
Abstract
Plants and animals use cell surface receptors to sense and interpret environmental signals. In legume symbiosis with nitrogen-fixing bacteria, the specific recognition of bacterial lipochitooligosaccharide (LCO) signals by single-pass transmembrane receptor kinases determines compatibility. Here, we determine the structural basis for LCO perception from the crystal structures of two lysin motif receptor ectodomains and identify a hydrophobic patch in the binding site essential for LCO recognition and symbiotic function. We show that the receptor monitors the composition of the amphiphilic LCO molecules and uses kinetic proofreading to control receptor activation and signaling specificity. We demonstrate engineering of the LCO binding site to fine-tune ligand selectivity and correct binding kinetics required for activation of symbiotic signaling in plants. Finally, the hydrophobic patch is found to be a conserved structural signature in this class of LCO receptors across legumes that can be used for in silico predictions. Our results provide insights into the mechanism of cell-surface receptor activation by kinetic proofreading of ligands and highlight the potential in receptor engineering to capture benefits in plant-microbe interactions.
Keywords: LysM receptors; kinetic proofreading; legume symbiosis; lipochitooligosaccharide signaling; receptor–ligand interaction.
Copyright © 2021 the Author(s). Published by PNAS.
Conflict of interest statement
Competing interest statement: K.G., Z.B., L.H.M., S.R., J.S., and K.R.A., are inventors on patent application 62/718,186 submitted by Aarhus University that covers LysM receptors: genetically altered LysM receptors with altered agonist specificity and affinity. M.L., D.L., S.R., J.S., and K.R.A., are inventors on patent application 63/027,151 submitted by Aarhus University that covers LysM receptor motifs.
Figures
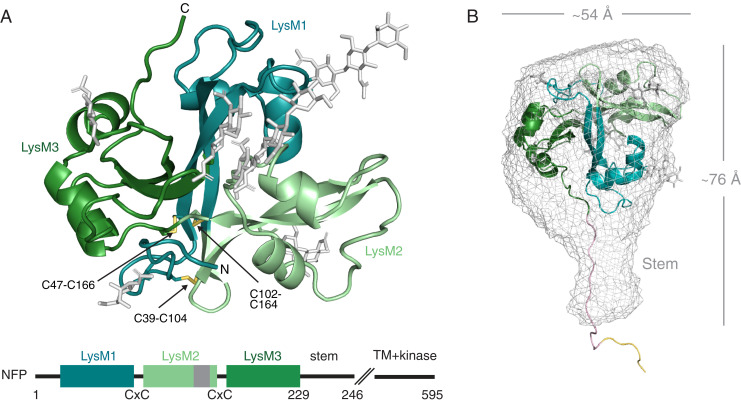
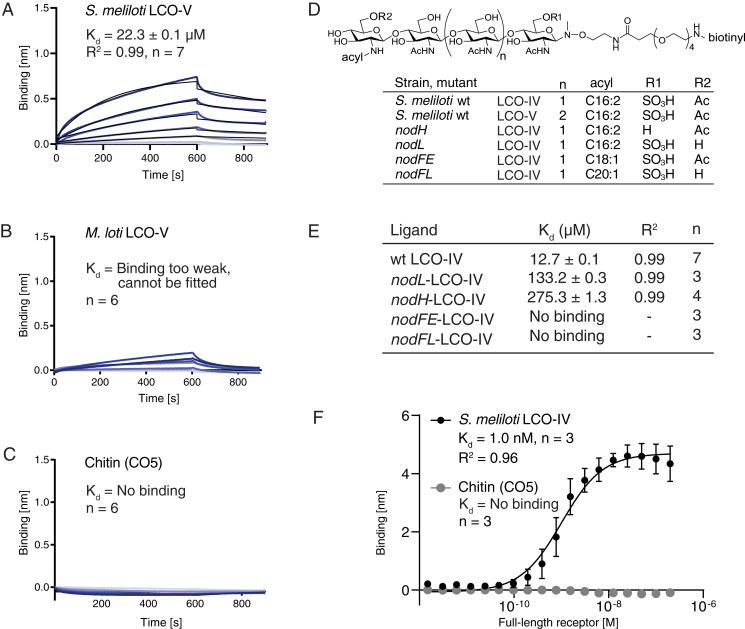
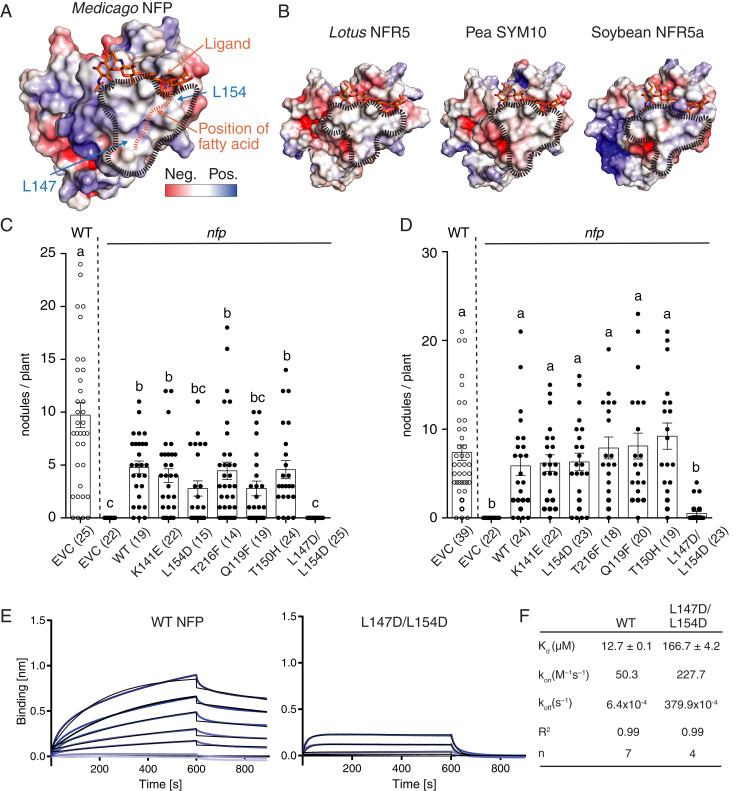
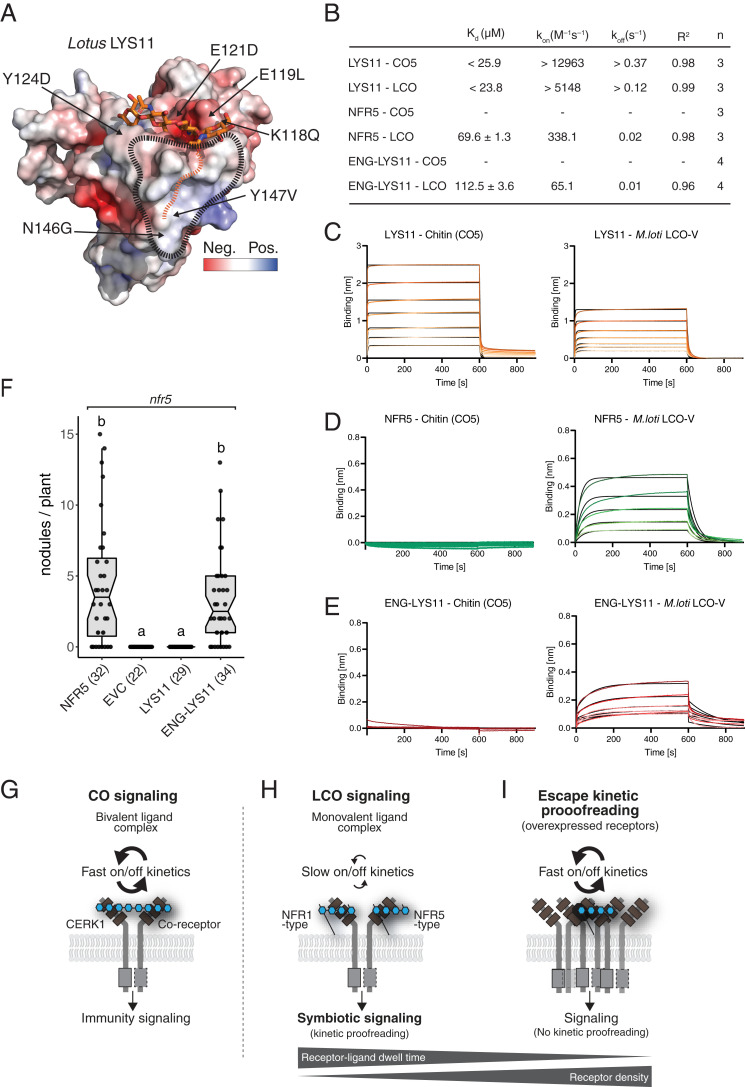
References
-
- D’Haeze W., Holsters M., Nod factor structures, responses, and perception during initiation of nodule development. Glycobiology 12, 79R–105R (2002). - PubMed
-
- Pueppke S. G., Broughton W. J., Rhizobium sp. strain NGR234 and R. fredii USDA257 share exceptionally broad, nested host ranges. Mol. Plant Microbe Interact. 12, 293–318 (1999). - PubMed
-
- Kelly S., Radutoiu S., Stougaard J., Legume LysM receptors mediate symbiotic and pathogenic signalling. Curr. Opin. Plant Biol. 39, 152–158 (2017). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
