

Genome and systems biology of Melilotus albus provides insights into coumarins biosynthesis
- PMID: 34717292
- PMCID: PMC8882801
- DOI: 10.1111/pbi.13742
Genome and systems biology of Melilotus albus provides insights into coumarins biosynthesis
Abstract
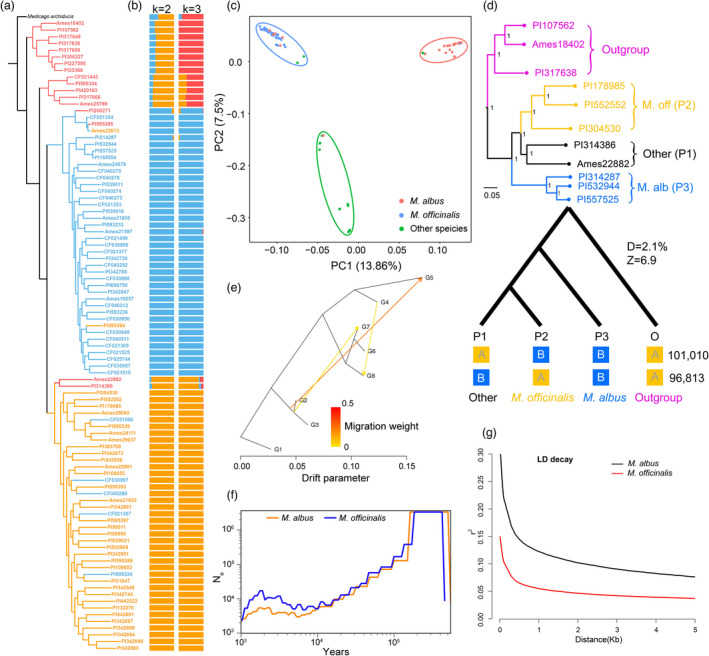
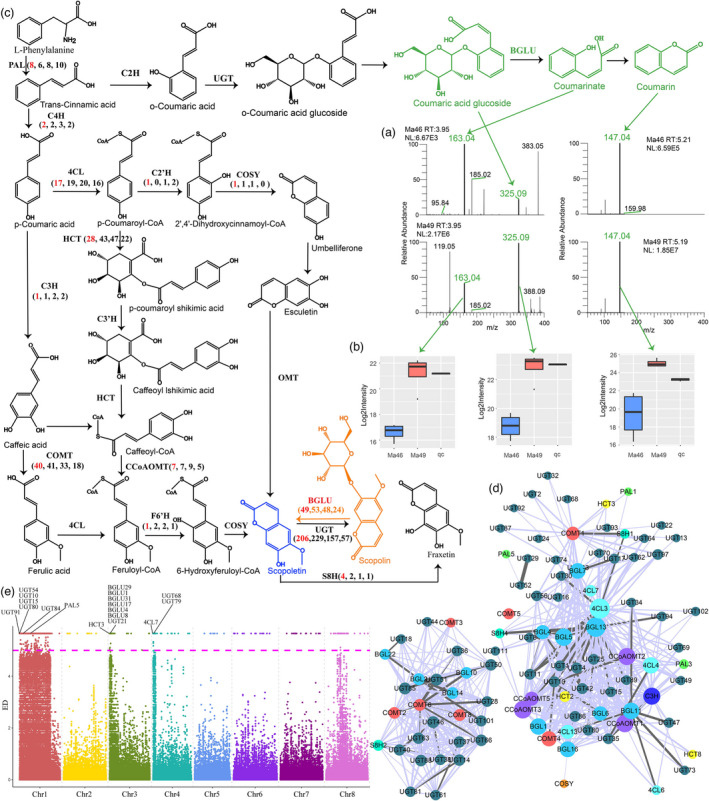
Melilotus species are used as green manure and rotation crops worldwide and contain abundant pharmacologically active coumarins. However, there is a paucity of information on its genome and coumarin production and function. Here, we reported a chromosome-scale assembly of Melilotus albus genome with 1.04 Gb in eight chromosomes, containing 71.42% repetitive elements. Long terminal repeat retrotransposon bursts coincided with declining of population sizes during the Quaternary glaciation. Resequencing of 94 accessions enabled insights into genetic diversity, population structure, and introgression. Melilotus officinalis had relatively larger genetic diversity than that of M. albus. The introgression existed between M. officinalis group and M. albus group, and gene flows was from M. albus to M. officinalis. Selection sweep analysis identified candidate genes associated with flower colour and coumarin biosynthesis. Combining genomics, BSA, transcriptomics, metabolomics, and biochemistry, we identified a β-glucosidase (BGLU) gene cluster contributing to coumarin biosynthesis. MaBGLU1 function was verified by overexpression in M. albus, heterologous expression in Escherichia coli, and substrate feeding, revealing its role in scopoletin (coumarin derivative) production and showing that nonsynonymous variation drives BGLU enzyme activity divergence in Melilotus. Our work will accelerate the understanding of biologically active coumarins and their biosynthetic pathways, and contribute to genomics-enabled Melilotus breeding.
Keywords: BGLU; coumarin biosynthesis; genome evolution; population genetics; sweet clover.
© 2021 The Authors. Plant Biotechnology Journal published by Society for Experimental Biology and The Association of Applied Biologists and John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures




References
-
- Ahn, Y.O. , Shimizu, B. , Sakata, K. , Gantulga, D. , Zhou, C. , Bevan, D.R. et al. (2010) Scopolin‐hydrolyzing beta‐glucosidases in roots of Arabidopsis . Plant Cell Physiol. 51, 132–143. - PubMed
-
- Barrett, J.C. , Fry, B. , Maller, J. and Daly, M.J. (2005) Haploview: analysis and visualization of LD and haplotype maps. Bioinformatics, 21, 263–265. - PubMed
-
- Bello, R.I. , Gomez‐Diaz, C. , Lopez‐Lluch, G. , Forthoffer, N. , Cordoba‐Pedregosa, M.C. , Navas, P. and Villalba, J.M. (2005) Dicoumarol relieves serum withdrawal‐induced G0/1 blockade in HL‐60 cells through a superoxide‐dependent mechanism. Biochem. Pharmacol. 69, 1613–1625. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
