

Role of the IgG4-related cholangitis autoantigen annexin A11 in cholangiocyte protection
- PMID: 34718050
- PMCID: PMC10804347
- DOI: 10.1016/j.jhep.2021.10.009
Role of the IgG4-related cholangitis autoantigen annexin A11 in cholangiocyte protection
Abstract
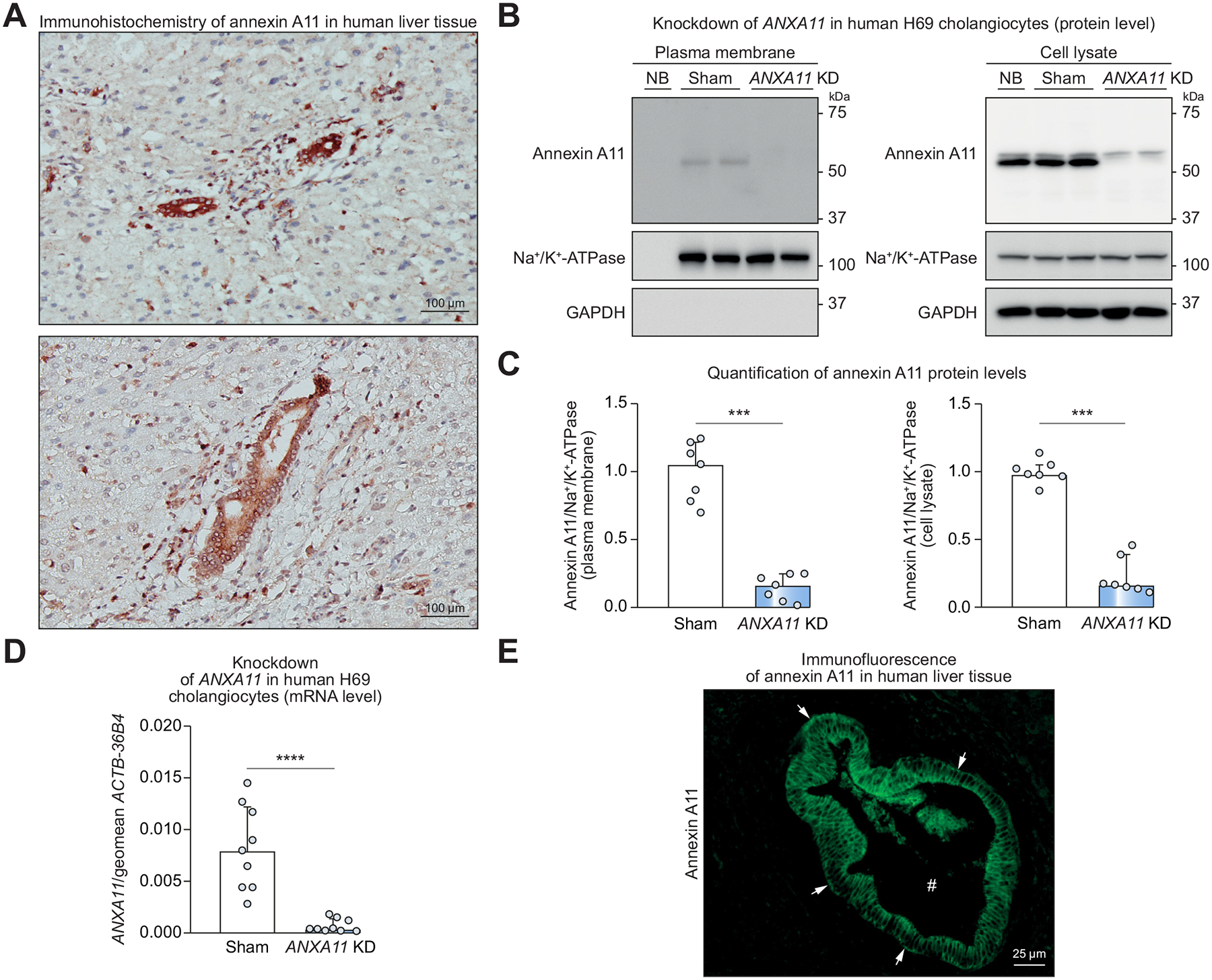
Background & aims: Annexin A11 was identified as autoantigen in IgG4-related cholangitis (IRC), a B-cell driven disease. Annexin A11 modulates calcium-dependent exocytosis, a crucial mechanism for insertion of proteins into their target membranes. Human cholangiocytes form an apical 'biliary bicarbonate umbrella' regarded as defense against harmful hydrophobic bile acid influx. The bicarbonate secretory machinery comprises the chloride/bicarbonate exchanger AE2 and the chloride channel ANO1. We aimed to investigate the expression and function of annexin A11 in human cholangiocytes and a potential role of IgG1/IgG4-mediated autoreactivity against annexin A11 in the pathogenesis of IRC.
Methods: Expression of annexin A11 in human liver was studied by immunohistochemistry and immunofluorescence. In human control and ANXA11 knockdown H69 cholangiocytes, intracellular pH, AE2 and ANO1 surface expression, and bile acid influx were examined using ratio microspectrofluorometry, cell surface biotinylation, and 22,23-3H-glycochenodeoxycholic acid permeation, respectively. The localization of annexin A11-mEmerald and ANO1-mCherry was investigated by live-cell microscopy in H69 cholangiocytes after incubation with IRC patient serum containing anti-annexin A11 IgG1/IgG4-autoantibodies or disease control serum.
Results: Annexin A11 was strongly expressed in human cholangiocytes, but not hepatocytes. Knockdown of ANXA11 led to reduced plasma membrane expression of ANO1, but not AE2, alkalization of intracellular pH and uncontrolled bile acid influx. High intracellular calcium conditions led to annexin A11 membrane shift and colocalization with ANO1. Incubation with IRC patient serum inhibited annexin A11 membrane shift and reduced ANO1 surface expression.
Conclusion: Cholangiocellular annexin A11 mediates apical membrane abundance of the chloride channel ANO1, thereby supporting biliary bicarbonate secretion. Insertion is inhibited by IRC patient serum containing anti-annexin A11 IgG1/IgG4-autoantibodies. Anti-annexin A11 autoantibodies may contribute to the pathogenesis of IRC by weakening the 'biliary bicarbonate umbrella'.
Lay summary: We previously identified annexin A11 as a specific autoantigen in immunoglobulin G4-related cholangitis (IRC), a B-cell driven disease affecting the bile ducts. Human cholangiocytes are protected against harmful hydrophobic bile acid influx by a defense mechanism referred to as the 'biliary bicarbonate umbrella'. We found that annexin A11 is required for the formation of a robust bicarbonate umbrella. Binding of patient-derived annexin A11 autoantibodies inhibits annexin A11 function, possibly contributing to bile duct damage by weakening the biliary bicarbonate umbrella in patients with IRC.
Keywords: IgG4-related systemic disease; annexin A11; anoctamin-1; autoimmunity; bicarbonate umbrella; cholangiopathy; membrane trafficking.
Copyright © 2021 The Author(s). Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare no conflict of interest with regard to this work. Please refer to the accompanying ICMJE disclosure forms for further details.
Figures





References
-
- Stone JH, Zen Y, Deshpande V. IgG4-related disease. N Engl J Med 2012;366:539–551. - PubMed
-
- Ghazale A, Chari ST, Zhang L, Smyrk TC, Takahashi N, Levy MJ, et al. Immunoglobulin G4-associated cholangitis: clinical profile and response to therapy. Gastroenterology 2008;134:706–715. - PubMed
-
- Maillette de Buy Wenniger LJ, Doorenspleet ME, Klarenbeek PL, Verheij J, Baas F, Elferink RP, et al. Immunoglobulin G4+ clones identified by next-generation sequencing dominate the B cell receptor repertoire in immunoglobulin G4 associated cholangitis. Hepatology 2013;57:2390–2398. - PubMed
-
- Doorenspleet ME, Hubers LM, Culver EL, Maillette de Buy Wenniger LJ, Klarenbeek PL, Chapman RW, et al. Immunoglobulin G4(+) B-cell receptor clones distinguish immunoglobulin G 4-related disease from primary sclerosing cholangitis and biliary/pancreatic malignancies. Hepatology 2016;64:501–507. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
