

Limited plasticity of anatomical and hydraulic traits in aspen trees under elevated CO2 and seasonal drought
- PMID: 34718790
- PMCID: PMC8774844
- DOI: 10.1093/plphys/kiab497
Limited plasticity of anatomical and hydraulic traits in aspen trees under elevated CO2 and seasonal drought
Abstract
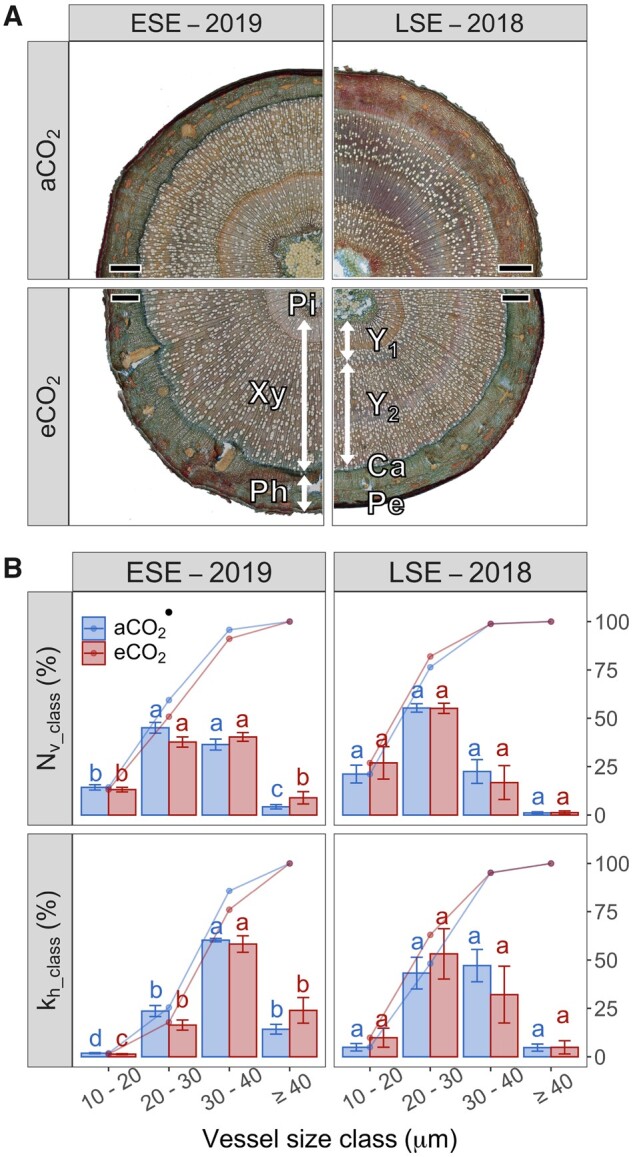
The timing of abiotic stress elicitors on wood formation largely affects xylem traits that determine xylem efficiency and vulnerability. Nonetheless, seasonal variability of elevated CO2 (eCO2) effects on tree functioning under drought remains largely unknown. To address this knowledge gap, 1-year-old aspen (Populus tremula L.) trees were grown under ambient (±445 ppm) and elevated (±700 ppm) CO2 and exposed to an early (spring/summer 2019) or late (summer/autumn 2018) season drought event. Stomatal conductance and stem shrinkage were monitored in vivo as xylem water potential decreased. Additional trees were harvested for characterization of wood anatomical traits and to determine vulnerability and desorption curves via bench dehydration. The abundance of narrow vessels decreased under eCO2 only during the early season. At this time, xylem vulnerability to embolism formation and hydraulic capacitance during severe drought increased under eCO2. Contrastingly, stomatal closure was delayed during the late season, while hydraulic vulnerability and capacitance remained unaffected under eCO2. Independently of the CO2 treatment, elastic, and inelastic water pools depleted simultaneously after 50% of complete stomatal closure. Our results suggest that the effect of eCO2 on drought physiology and wood traits are small and variable during the growing season and question a sequential capacitive water release from elastic and inelastic pools as drought proceeds.
© American Society of Plant Biologists 2021. All rights reserved. For permissions, please email: journals.permissions@oup.com.
Figures




References
-
- Adams HD, Zeppel MJB, Anderegg WRL, Hartmann H, Landhäusser SM, Tissue DT, Huxman TE, Hudson PJ, Franz TE, Allen CD, et al. (2017) A multi-species synthesis of physiological mechanisms in drought-induced tree mortality. Nat Ecol Evol 1: 1285–1291 - PubMed
-
- Ainsworth EA, Rogers A (2007) The response of photosynthesis and stomatal conductance to rising [CO 2]: mechanisms and environmental interactions. Plant Cell Environ 30: 258–270 - PubMed
-
- Alder NN, Pockman WT, Sperry JS, Nuismer S (1997) Use of centrifugal force in the study of xylem cavitation. J Exp Bot 48: 665–674
-
- Allen RG, Pereira LS, Raes D, Smith M (1998) Crop evapotranspiration — guidelines for computing crop water requirement. FAO Irrig Drain Pap 56. Food and Agriculture Organization, Rome
-
- Atwell BJ, Henery ML, Whitehead D (2003) Sapwood development in Pinus radiata trees grown for three years at ambient and elevated carbon dioxide partial pressures. Tree Physiol 23: 13–21 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
