

Iron-Dependent Autophagic Cell Death Induced by Radiation in MDA-MB-231 Breast Cancer Cells
- PMID: 34722507
- PMCID: PMC8551752
- DOI: 10.3389/fcell.2021.723801
Iron-Dependent Autophagic Cell Death Induced by Radiation in MDA-MB-231 Breast Cancer Cells
Abstract
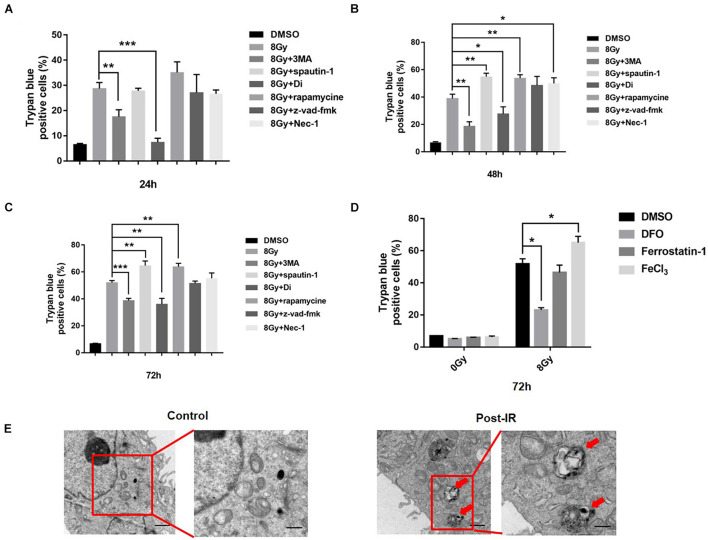
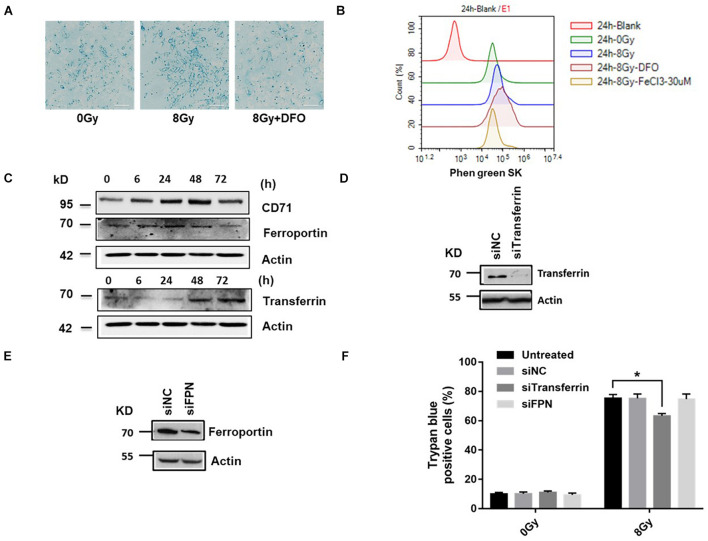
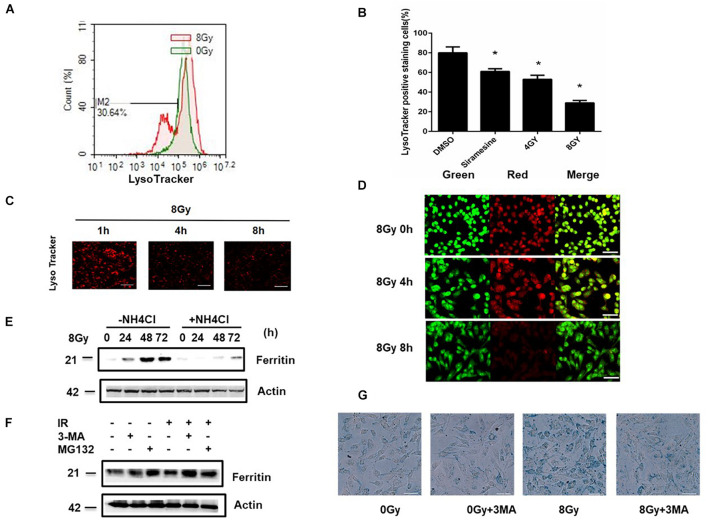
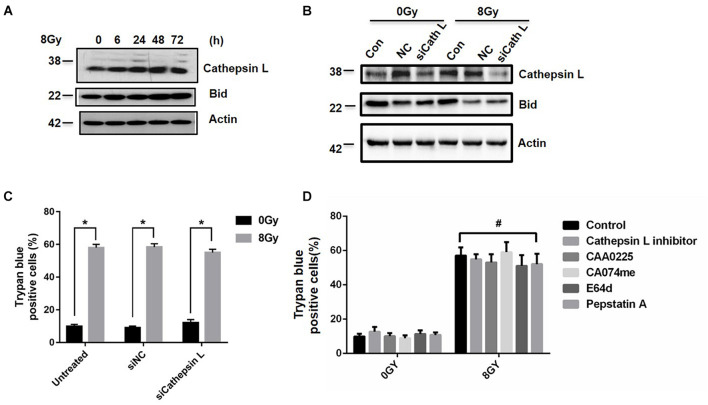
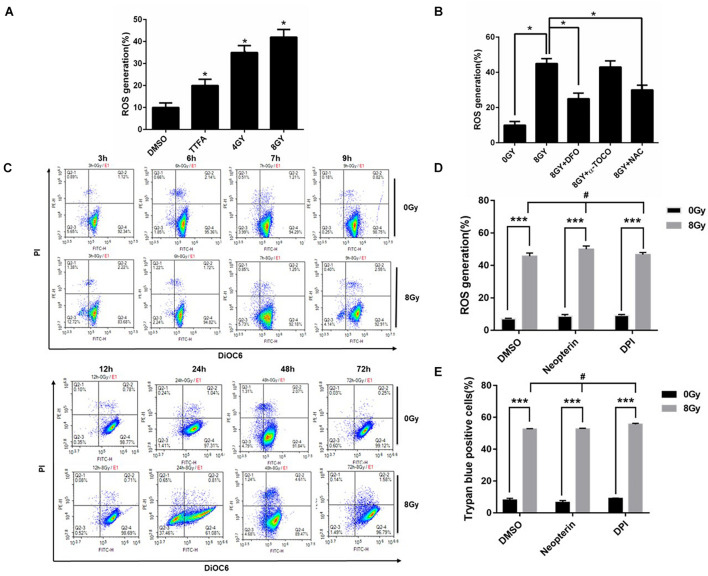
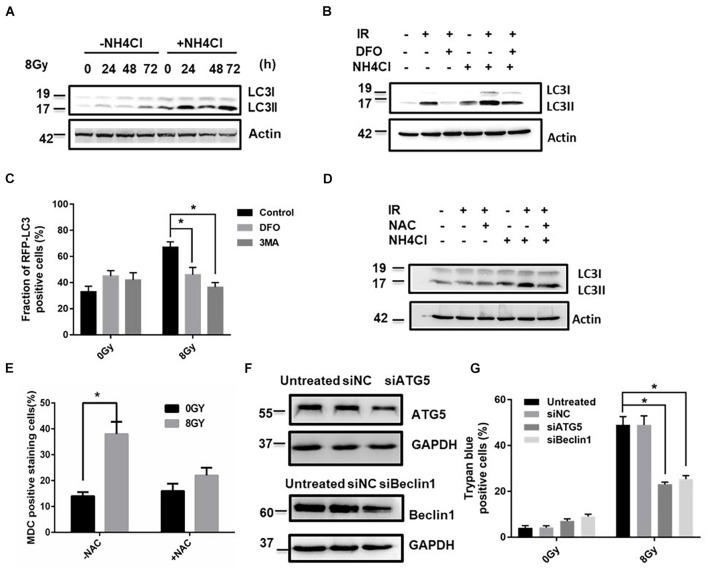
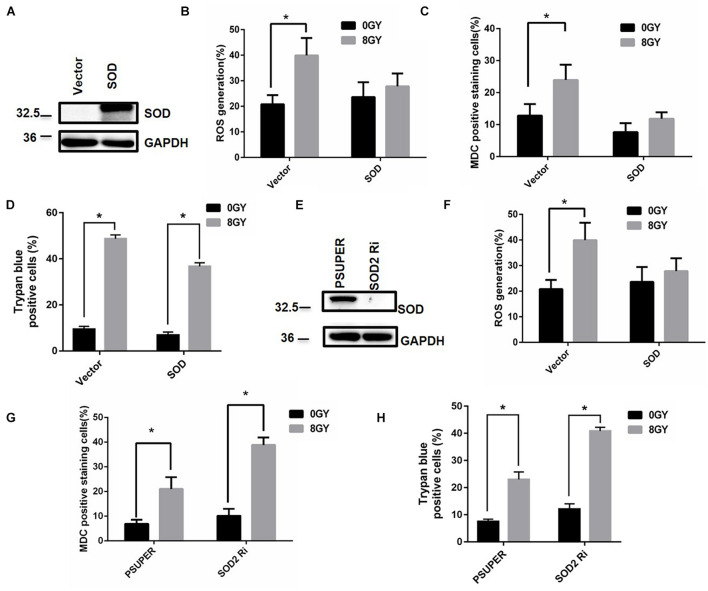
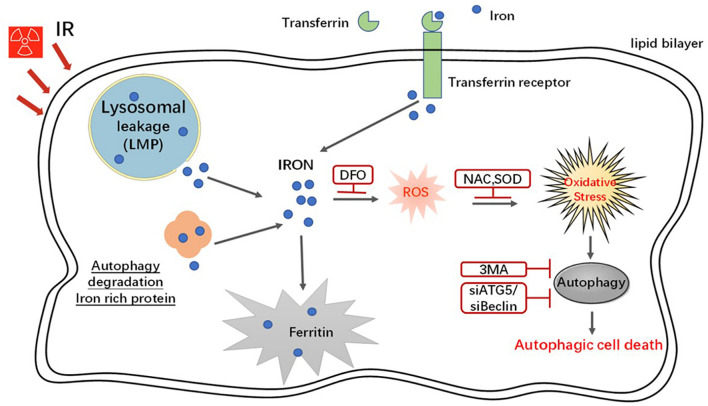
In radiation oncology, ionizing radiation is used to kill cancer cells, in other words, the induction of different types of cell death. To investigate this cellular death and the associated iron accumulation, the transfer, release, and participation of iron after radiation treatment was analyzed. We found that radiation-induced cell death varied in different breast cancer cells and autophagy was induced in MDA-MB-231 and BT549 cells (triple negative breast cancer cell line) rather than in MCF-7 and zr-75 cells. Iron chelator deferoxamine (DFO), the autophagy inhibitor 3MA, silencing of the autophagy-related genes ATG5, and Beclin 1 could decrease radiation induced cell death in MDA-MB-231 cells, while inhibitors of apoptosis such as Z-VAD-FMK, ferroptosis inhibitor ferrostatin-1 (Fer-1), and necroptosis inhibitor Necrostatin-1 showed no change. This suggests the occurrence of autophagic cell death. Furthermore, we found that iron accumulation and iron regulatory proteins, including transferrin (Tf), transferrin receptor (CD71), and Ferritin (FTH), increased after radiation treatment, and the silencing of transferrin decreased radiation-induced cell death. In addition, radiation increased lysosomal membrane permeabilization (LMP) and the release of lysosomal iron and cathepsins, while cathepsins silencing failed to change cell viability. Radiation-induced iron accumulation increased Reactive oxygen species (ROS) generation via the Fenton reaction and increased autophagy in a time-dependent manner. DFO, N-acetylcysteine (NAC), and overexpression of superoxide dismutase 2 (SOD2) decreased ROS generation, autophagy, and cell death. To summarize, for the first time, we found that radiation-induced autophagic cell death was iron-dependent in breast cancer MDA-MB-231 cells. These results provide new insights into the cell death process of cancers and might conduce to the development and application of novel therapeutic strategies for patients with apoptosis-resistant breast cancer.
Keywords: autophagy; breast cancer; iron; lysosome membrane permeabilization (LMP); radiation; reactive oxygen species (ROS).
Copyright © 2021 Ma, Fu, Liu, Liu, Feng, Jiang, Liu, Liu, Liang, Li, Tian, Hu, Bai, Liang and Liu.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
-
- Desouky O., Ding N., Zhou G. (2015). Targeted and non-targeted effects of ionizing radiation. J. Rad. Res. Appl. Sci. 8 247–254. 10.1016/j.jrras.2015.03.003 - DOI
LinkOut - more resources
Full Text Sources
Miscellaneous