

Understanding the Logistics for the Distribution of Heme in Cells
- PMID: 34723258
- PMCID: PMC8549057
- DOI: 10.1021/jacsau.1c00288
Understanding the Logistics for the Distribution of Heme in Cells
Abstract
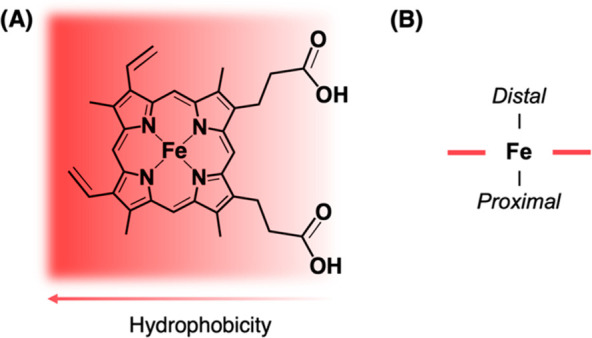
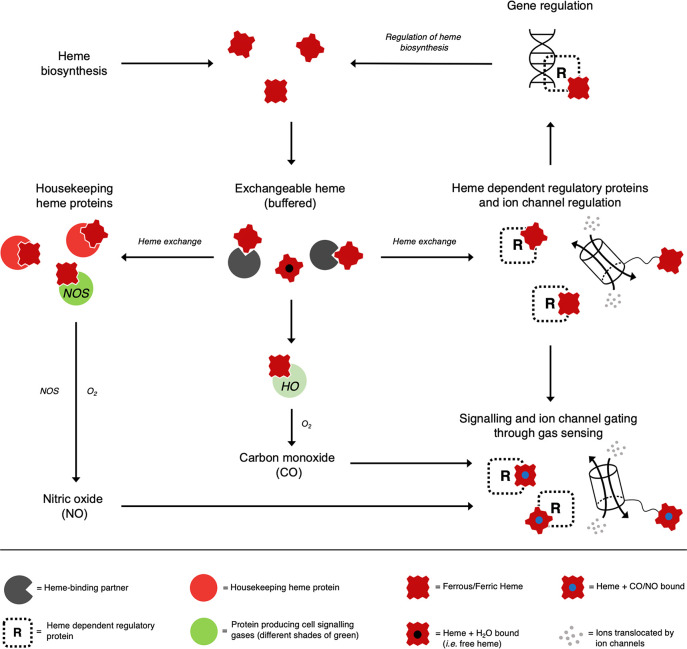
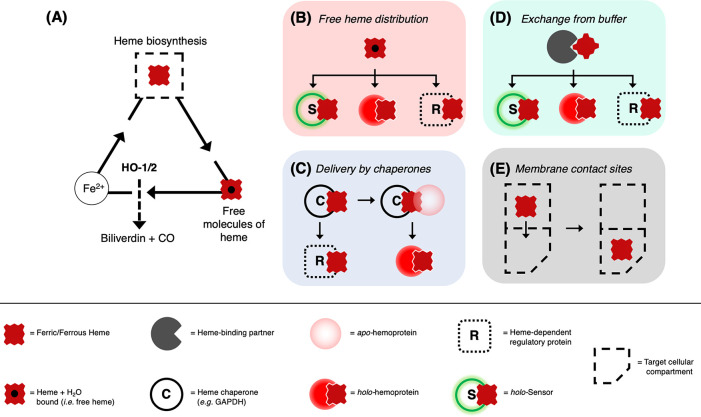
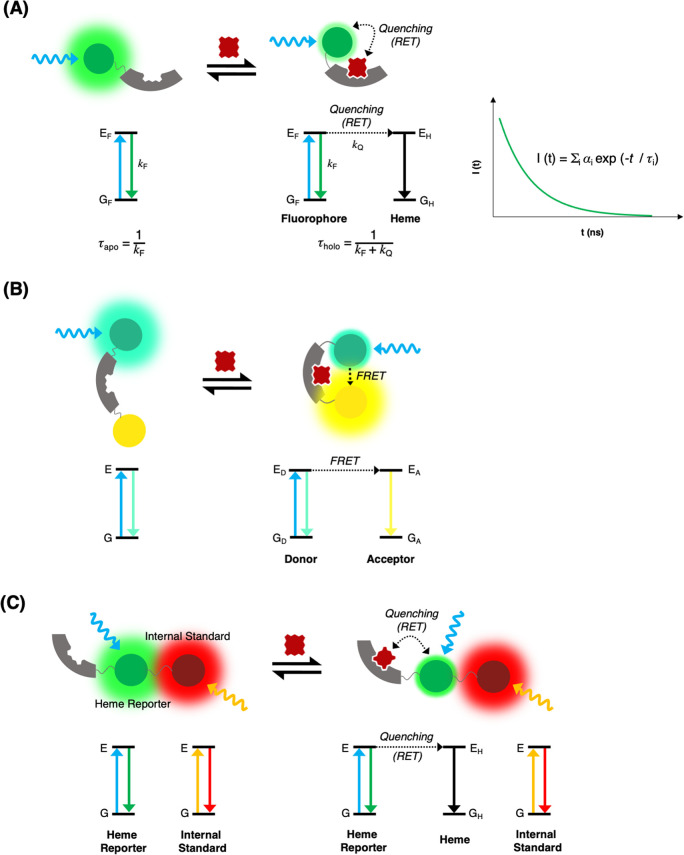
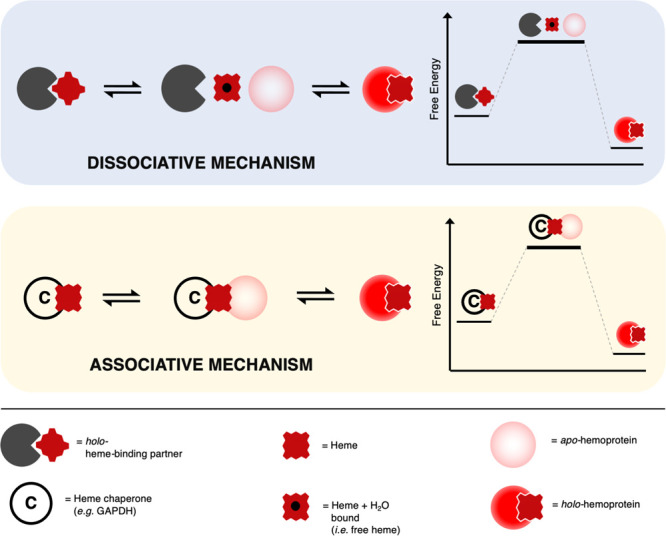
Heme is essential for the survival of virtually all living systems-from bacteria, fungi, and yeast, through plants to animals. No eukaryote has been identified that can survive without heme. There are thousands of different proteins that require heme in order to function properly, and these are responsible for processes such as oxygen transport, electron transfer, oxidative stress response, respiration, and catalysis. Further to this, in the past few years, heme has been shown to have an important regulatory role in cells, in processes such as transcription, regulation of the circadian clock, and the gating of ion channels. To act in a regulatory capacity, heme needs to move from its place of synthesis (in mitochondria) to other locations in cells. But while there is detailed information on how the heme lifecycle begins (heme synthesis), and how it ends (heme degradation), what happens in between is largely a mystery. Here we summarize recent information on the quantification of heme in cells, and we present a discussion of a mechanistic framework that could meet the logistical challenge of heme distribution.
© 2021 The Authors. Published by American Chemical Society.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
References
-
- Moss G. P.; Smith P. A. S.; Tavernier D. Glossary of class names of organic compounds and reactivity intermediates based on structure (IUPAC Recommendations 1995). Pure Appl. Chem. 1995, 67 (8–9), 1307–1375. 10.1351/pac199567081307. - DOI
Publication types
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous