

Population genomic analysis of Aegilops tauschii identifies targets for bread wheat improvement
- PMID: 34725503
- PMCID: PMC8926922
- DOI: 10.1038/s41587-021-01058-4
Population genomic analysis of Aegilops tauschii identifies targets for bread wheat improvement
Abstract
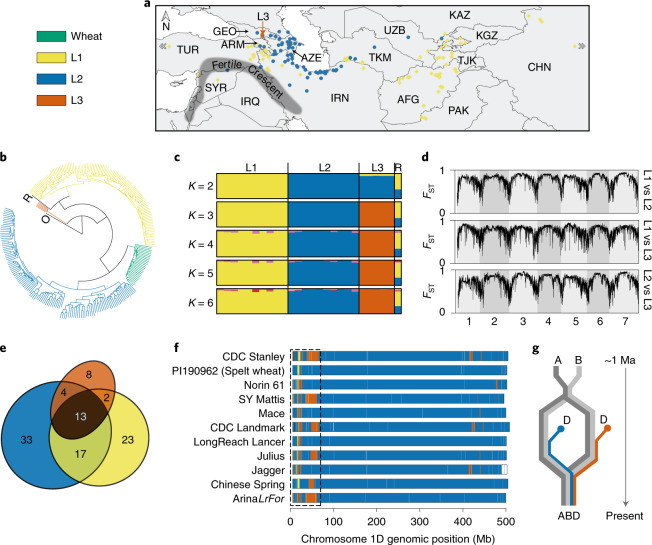
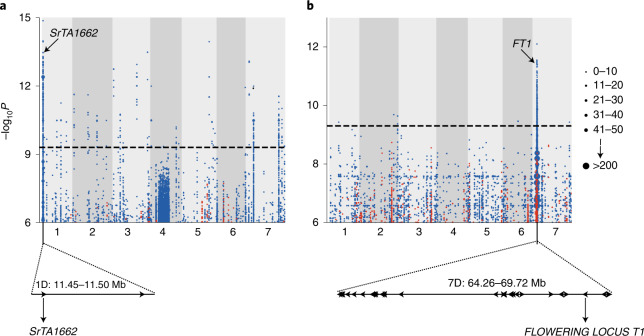
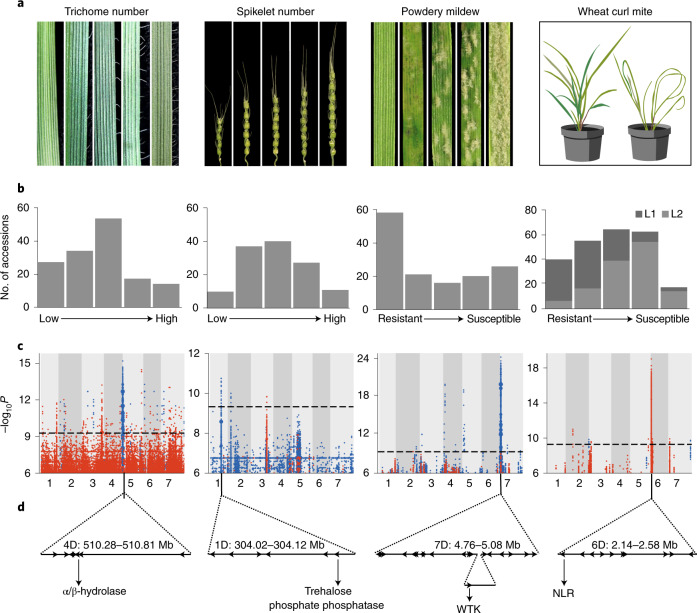
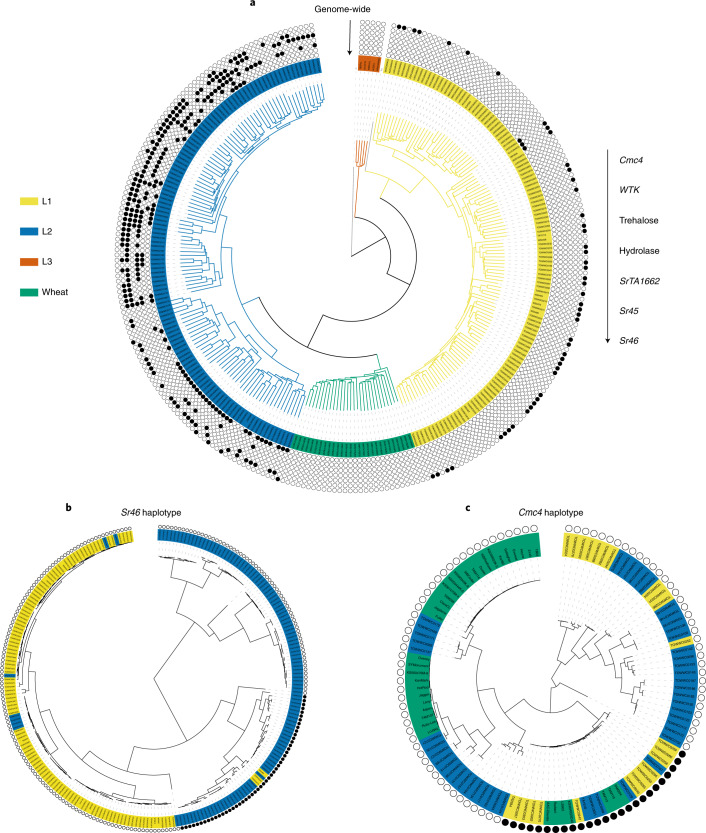
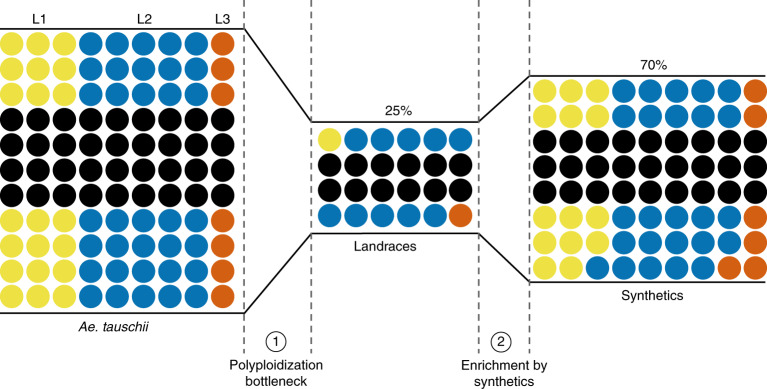
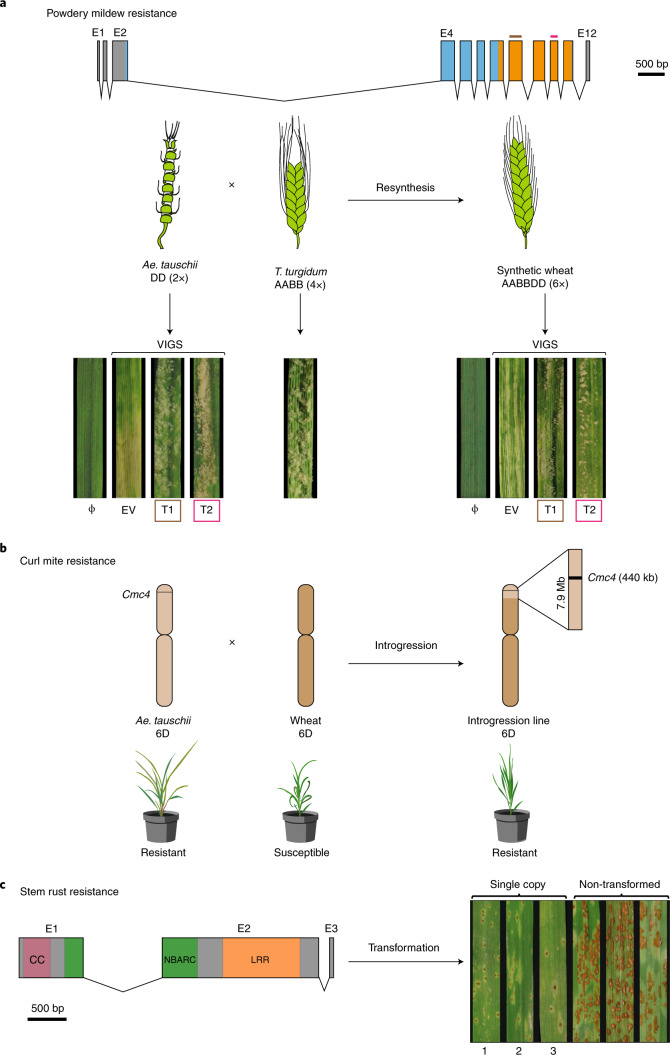
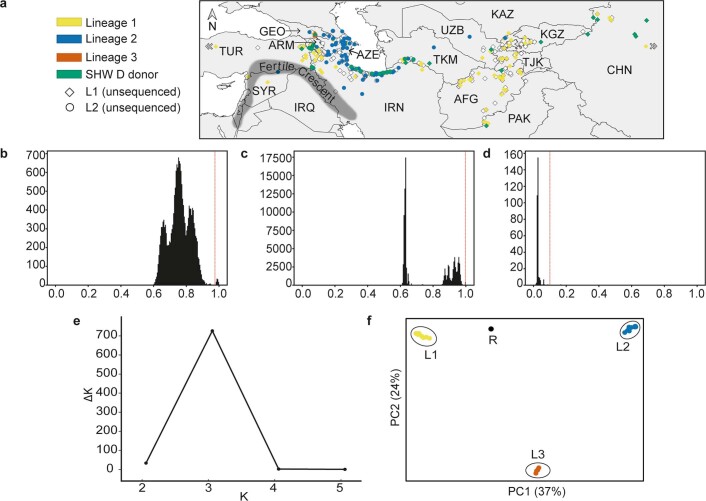
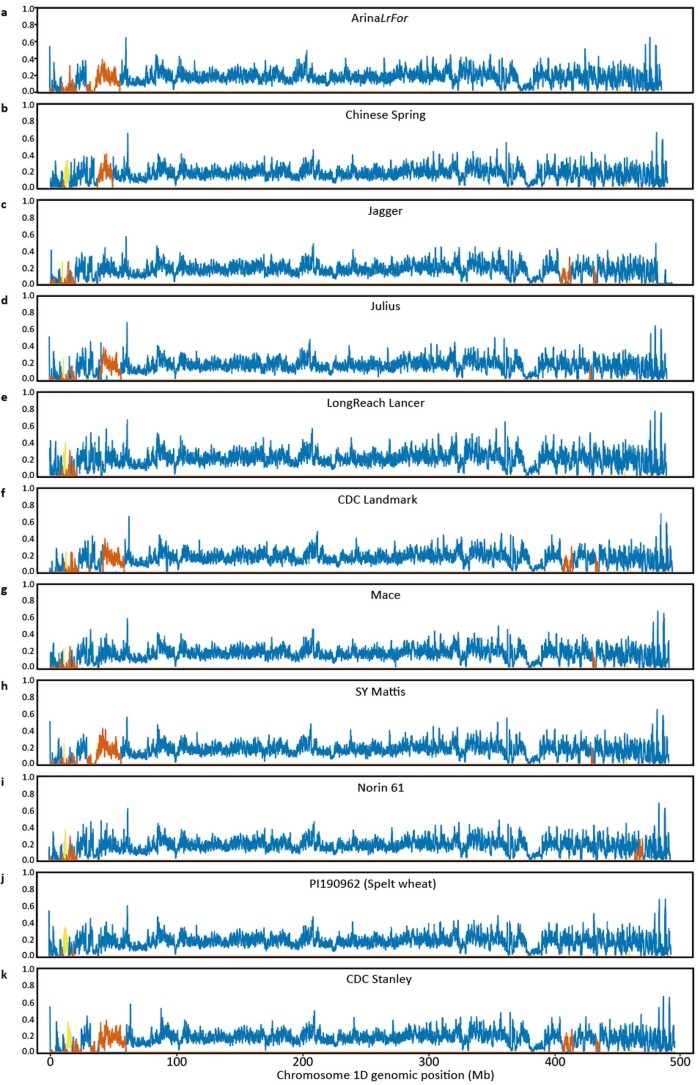
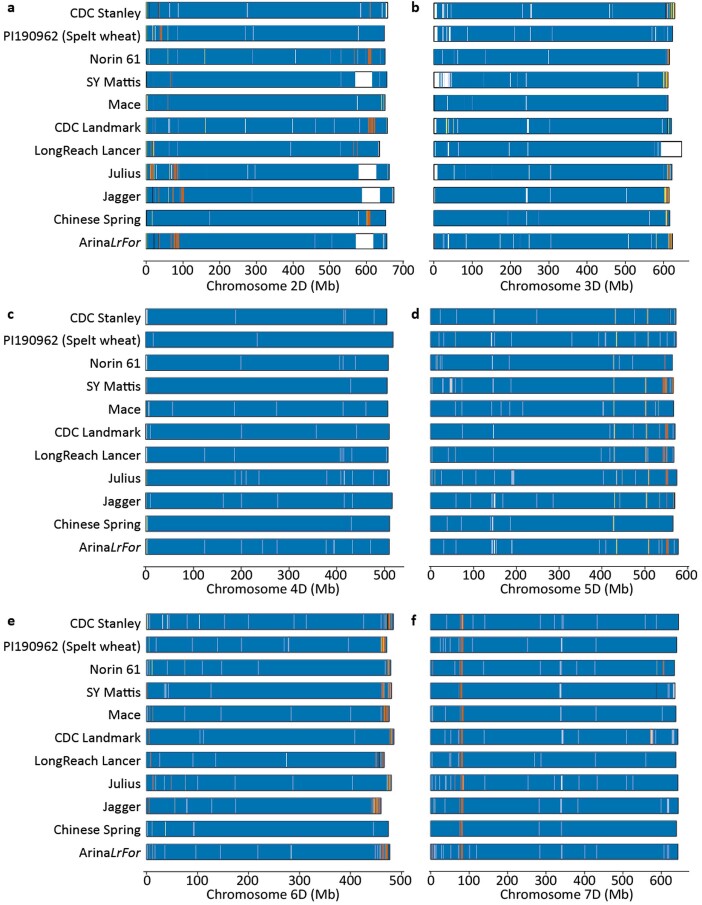
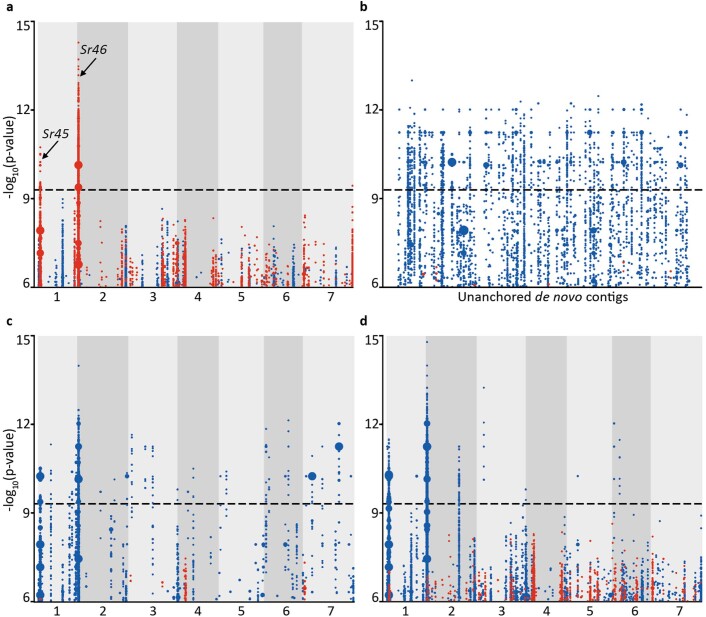
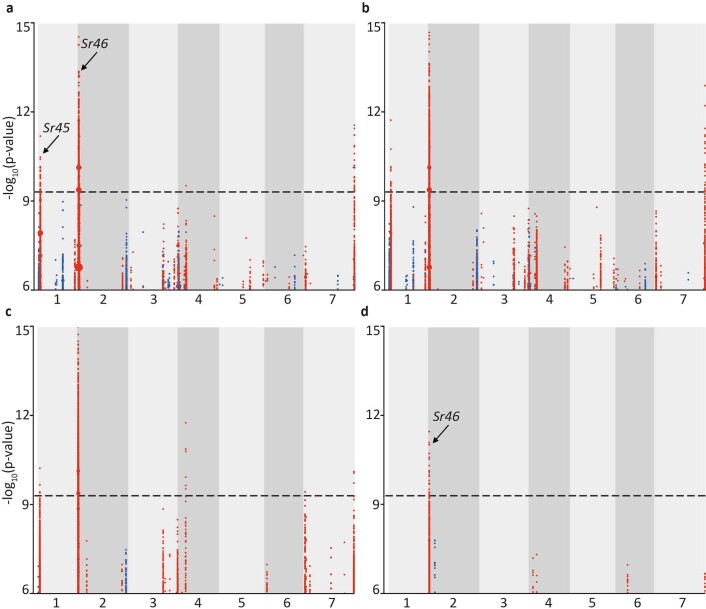
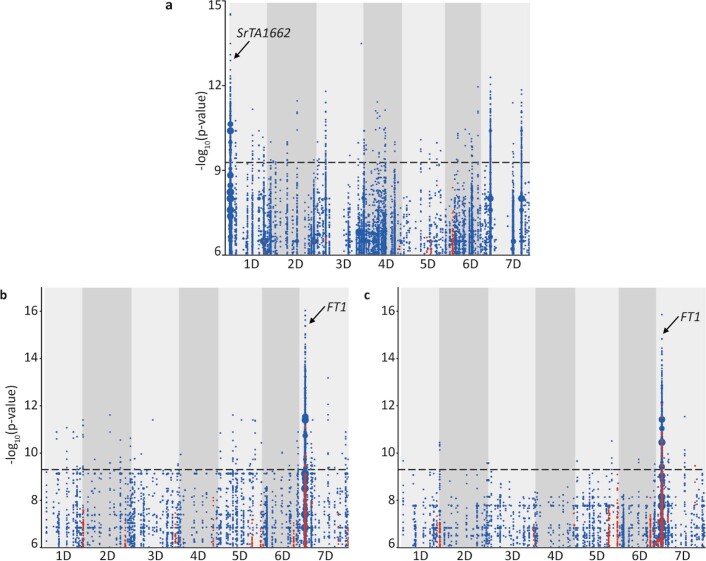
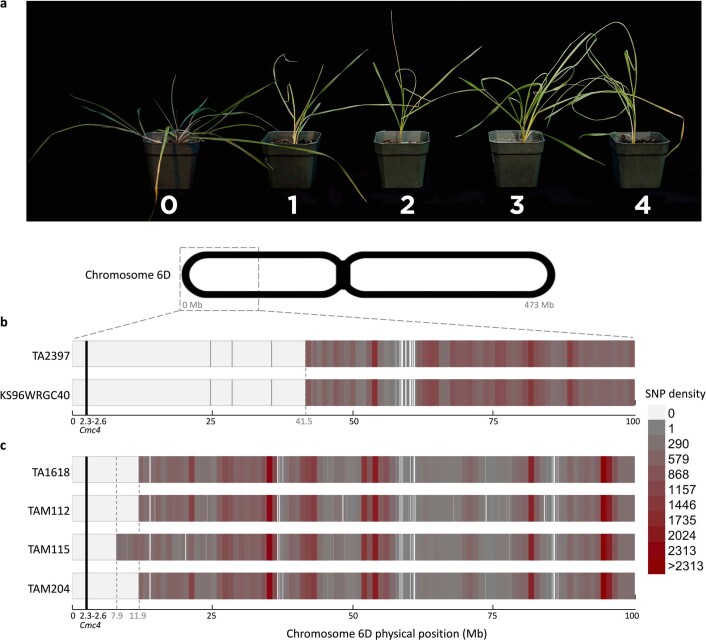
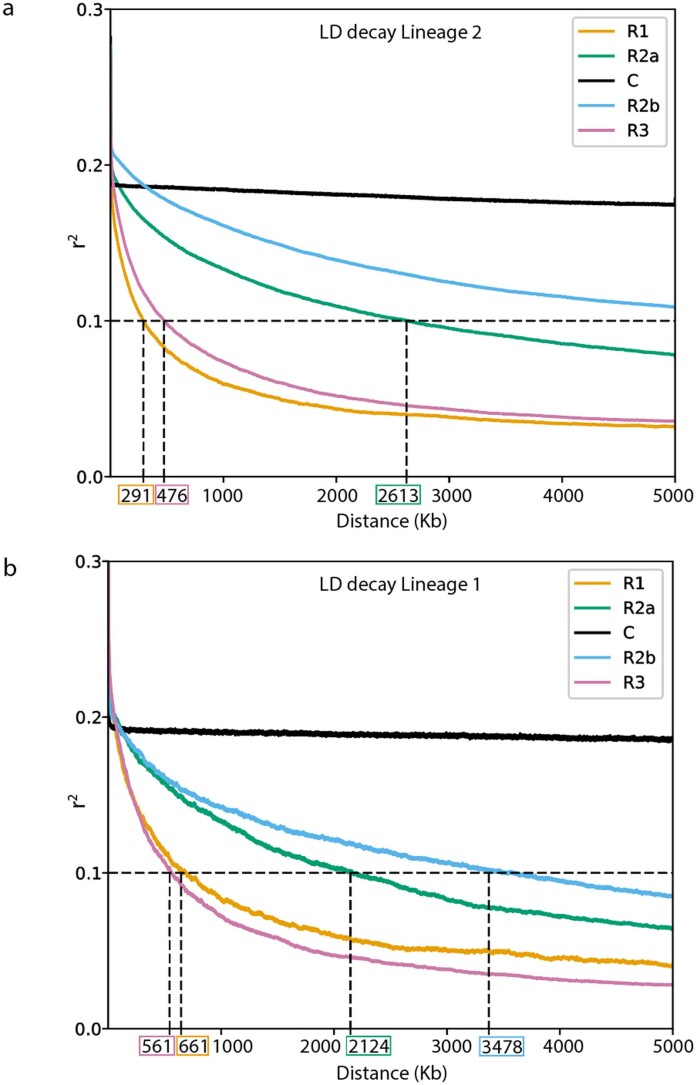
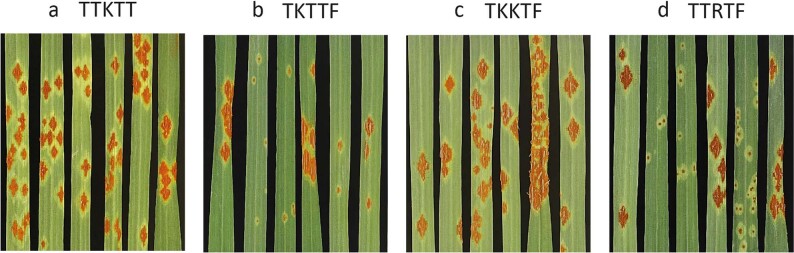
Aegilops tauschii, the diploid wild progenitor of the D subgenome of bread wheat, is a reservoir of genetic diversity for improving bread wheat performance and environmental resilience. Here we sequenced 242 Ae. tauschii accessions and compared them to the wheat D subgenome to characterize genomic diversity. We found that a rare lineage of Ae. tauschii geographically restricted to present-day Georgia contributed to the wheat D subgenome in the independent hybridizations that gave rise to modern bread wheat. Through k-mer-based association mapping, we identified discrete genomic regions with candidate genes for disease and pest resistance and demonstrated their functional transfer into wheat by transgenesis and wide crossing, including the generation of a library of hexaploids incorporating diverse Ae. tauschii genomes. Exploiting the genomic diversity of the Ae. tauschii ancestral diploid genome permits rapid trait discovery and functional genetic validation in a hexaploid background amenable to breeding.
© 2021. The Author(s).
Conflict of interest statement
K.G. and B.B.H.W. are inventors on UK patent application PC931335GB, and S. Arora, B. Steuernagel and B.B.H.W. are inventors on PCT/US2019/013430; these patents are based on part of the work presented here. The remaining authors declare no competing interests.
Figures

References
-
- Pont C, et al. Tracing the ancestry of modern bread wheats. Nat. Genet. 2019;51:905–911. - PubMed
-
- Marcussen T, et al. Ancient hybridizations among the ancestral genomes of bread wheat. Science. 2014;345:1250092. - PubMed
-
- Zohary, D., Hopf, M. & Weiss, E. Domestication of Plants in the Old World: The Origin and Spread of Domesticated Plants in Southwest Asia, Europe, and the Mediterranean Basin 4th edn (Oxford Scholarship Online, 2012).
Publication types
MeSH terms
Grants and funding
- BB/P016855/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/N019113/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BBS/E/J/000PR8000/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BBS/E/T/000PR9817/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BBS/E/T/000PR9814/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
